![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 47) Ano 2017. Pág. 21
Miguel Pesch TRAMONTINI 1; Paulo César FLÔRES Junior 2; Antonio Rioyei HIGA 3; Giovana Bomfim de ALCANTARA 4
Recebido: 29/05/2017 • Aprovado: 30/06/2017
RESUMO: O objetivo deste trabalho foi avaliar a influência do ácido giberélico (0, 250, 500 e 750 mg L-1) e de métodos de produção de sementes (área de produção de sementes e formigueiro) na germinação e no crescimento de Acacia mearnsii. O delineamento foi em blocos casualizados em esquema fatorial. As sementes de formigueiro apresentaram melhor germinação, índice de velocidade de germinação e comprimento de raiz. O ácido giberélico (750 mg L-1) proporcionou maior comprimento de parte aérea e hipocótilo. |
ABSTRACT: The objective of this study was to evaluate the effects of gibberellic acid (0, 250, 500 e 750 mg L-1) and seeds production methods (area of seed production and ant-nest) on germination and growth of Acacia mearnsii. A randomized complete block design was adopted. Ant-nest seed show higher percentage of germination, germination speed index and root length. The gibberellic acid (750 mg L-1) showed better response on length aerial part and hypocotyl. |

Acácia negra (Acacia mearnsii DE WILDEMANN) foi introduzida no Brasil no início do século XX e os seus primeiros plantios comerciais tiveram início no ano de 1928. Mas foi a partir da década de 1980 que a espécie começou a ser trabalhada dentro de programas de melhoramento, por meio da criação de Áreas de Produção de Sementes (APS) visando sementes de qualidade genética para produtividade.
Atualmente no Rio Grande do Sul há 101.439 ha de plantações de acácia negra (AGEFLOR, 2014), e o gênero acácia possui 168.827 ha no Brasil (IBA, 2016). As áreas de plantio da espécie são ainda hoje prioritariamente originárias de mudas seminais (entre 97 e 98%), o que apresenta uma necessidade de estudos que priorizem a maximização da germinação de sementes para atender a demanda de mudas da espécie. É importante ressaltar que estudos vem sendo realizados para otimizar e solucionar processos de clonagem, tanto por técnicas de miniestacas e micropropagação (Disarz, Martins-Corder, 2009; Engel et al., 2017).
A espécie apresenta características multifuncionais tornando-a muito interessante tanto do ponto de vista de produção quanto ecológico. É uma espécie que apresenta um ciclo de rotação semelhante ao do eucalipto, gênero mais plantado no Brasil, variando de 5,5 a 7 anos (Riegel et al., 2008). Dentro do setor produtivo, a importância da espécie se apresenta em uma gama de subprodutos como extração de taninos presentes na casca, sua madeira apresenta alto poder calorífico, sendo priorizada para energia, podendo ser utilizada também na indústria farmacêutica, floculantes para limpeza de recursos hídricos, entre outros usos. Dentre as funções ecológicas, a espécie apresenta capacidade de fixação de nitrogênio de aproximadamente 200 Kg ha-1 ano-1 (Auer, Silva, 1992) com isso atua também na recuperação de solos degradados bem como pode ser utilizada em consórcios com outras culturas.
A área de produção de sementes consiste em utilizar uma área com um povoamento já estabelecido, onde há conservação genética, e efetuada a seleção de árvores fenotipicamente superiores, com o desbaste das demais. Este método caracteriza-se por produzir material genético superior em curto prazo e baixo custo (Pires et al., 2011, p. 229). Outra procedência de sementes utilizada para a espécie é a coleta diretamente de formigueiros que se encontram dentro das áreas produtoras de sementes e talhões comerciais no Rio Grande do Sul.
As sementes de acácia negra são dispersas pelas formigas, em um processo conhecido por mirmecocoria (Handel, Beattie, 1990; Fokuhl, Heinze, Poschlod, 2012), e levadas até os formigueiros. As formigas do tipo cortadeiras tem interesse nas sementes de acácia negra devido ao arilo (Mochiutti, Higa, Simon, 2007), uma camada rica em gordura, depositada no interior do formigueiro para crescimento do fungo que servirá como alimento. Após a retirada do arilo, a semente já não é mais interessante, sendo depositada no local chamado de “lixeira”, ao lado de fora do formigueiro (Mora, 2002).
O ácido giberélico (GA) ou giberelina regula um amplo espectro de processos de desenvolvimento da planta, dentre os quais destacam-se a germinação de sementes, expansão foliar, alongamento do caule, florescimento e desenvolvimento de frutos e sementes (Gou et al., 2010). A giberelina tem sido utilizada com êxito em testes de germinação e produção de mudas em diversificadas espécies vegetais, tais como: Annona cherimola Mill. x A. squamosa L. (Oliveira et al., 2010), Campomanesia adamantium (Scalon et al., 2009) e Inga vera (Stein et al., 2007).
O objetivo deste trabalho foi verificar a influência dos métodos de produção de sementes de acácia negra e de concentrações do ácido giberélico na germinação, índice de velocidade de germinação (IVG), comprimento da raiz, comprimento da parte aérea e do hipocótilo.
O trabalho foi conduzido no Laboratório de Genética e Melhoramento Florestal (LAMEF), utilizando dois métodos de produção: área de produção de sementes (APS) e formigueiro. As sementes foram coletadas de plantações comerciais do Rio Grande do Sul em 2009 e armazenadas a 4-5ºC. Inicialmente foi realizada uma pré-seleção das sementes, as quais foram imersas em água e as que boiaram foram consideradas sementes chochas e portanto, descartadas. Posteriormente foi realizada a superação da dormência deixando as sementes imersas em água à cerca de 100ºC durante 1 minuto e da lavagem em água corrente durante 1 minuto (Martins-Corder, Borges Junior, 1999).
Após a superação da dormência as sementes foram submetidas à desinfestação, com imersão em álcool 70% por 1 minuto, hipoclorito de sódio 2% por 2 minutos e triplo enxágue em água autoclavada. Posteriormente em câmara de fluxo laminar, as sementes permaneceram em solução de ácido giberélico por 12 hs sem agitação nas seguintes concentrações: 0, 250, 500 e 750 mg L-1.
As sementes foram mantidas em caixas de acrílico do tipo Gearboxâ, de 11 x 11 cm, contendo papel filtro com lâmina d’água e foram armazenados em sala de crescimento, com temperatura de 25ºC e fotoperíodo de 16h luz. A análise de germinação foi realizada durante o período de 22 dias e as medições no 22º dia utilizando papel milimetrado e em câmara de fluxo laminar. Segundo a RAS (2009) este tempo de avaliação da germinação é de no mínimo 21 dias. Foram avaliadas as seguintes variáveis: porcentagem de germinação, índice de velocidade de germinação (IVG), comprimento da raiz (mm), comprimento da parte aérea (mm) e comprimento do hipocótilo (mm).
O delineamento experimental utilizado foi em blocos ao acaso em esquema fatorial 4x2 (tratamento de GA x métodos de produção), com seis repetições, sendo cada unidade experimental constituída por 20 sementes totalizando 960 sementes. Os dados obtidos foram inicialmente submetidos ao teste de normalidade (Shapiro-Wilk) e posteriormente à análise de variância e teste de Tukey a 5% de probabilidade utilizando o programa estatístico Assistat® versão 7.7, 2017 (Silva, Azevedo, 2016).
De acordo com os resultados da análise de variância observou-se que a interação entre os fatores concentrações de giberelina e métodos de produção de sementes não foi estatisticamente significativa para nenhuma das variáveis analisadas, indicando a independência dos fatores testados.
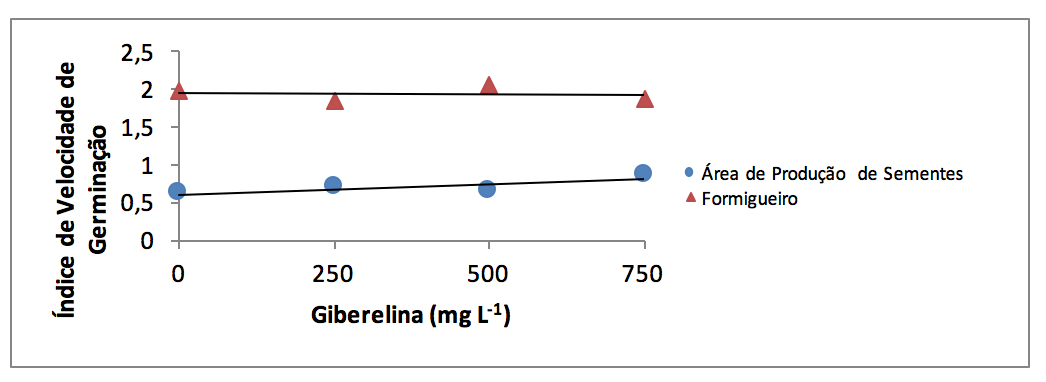
Os tratamentos com ácido giberélico não afetaram a germinação (Tabela 1), o índice de velocidade de germinação (IVG) (Gráfico 1) e o comprimento da raiz (Tabela 2), não havendo diferença estatística entre as concentrações de GA testadas para essas variáveis. Entretanto, para as variáveis comprimento da parte aérea (Tabela 3) e do hipocótilo (Tabela 4) foi verificada diferença estatística entre as concentrações de GA testadas. Com sementes de macadâmia utilizando GA também não foi evidenciada influência do regulador na germinação de sementes (Dalastra et al., 2010). Estes resultados podem ocorrer devido ao contato direto do embrião com umidade excessiva proporcionada pela solução de GA, antes de desencadear o processo germinativo, podendo ser prejudicial a germinação das sementes. O uso em demasia de embebição da semente pode ocasionar a liberação de solutos, reduzindo dessa forma o período disponível para que as membranas possam se reorganizar, ocorrendo então uma diminuição na germinação (Malavasi, 1988). A possível liberação de solutos ocasionada pelo excesso de embebição da semente pode ter prejudicado a ação da enzima a amilase o que não promoveu uma maior germinação no presente trabalho. O ácido giberélico atua na produção da enzima a amilase, propiciando a degradação do amido e consequentemente promovendo uma germinação mais rápida e uniforme (Flores et al. 2002).

Houve diferença estatística entre os dois métodos de produção de sementes para as variáveis porcentagem de germinação de sementes (Tabela 1), IVG (Gráfico 1) e comprimento de raízes (Tabela 2). O método de produção em formigueiro apresentou melhor resposta frente ao método em área de produção de sementes (APS). Esta diferença ocorre provavelmente devido à forma como a coleta das sementes é realizada em cada método. Na APS, para que seja economicamente viável, as sementes são coletadas ao mesmo tempo, isto faz com que muitas sementes não apresentem maturidade fisiológica no momento da coleta, o que pode ser visualizado pelas baixas porcentagens de germinação obtidas (37,5 a 46,67%) (Tabela 1). Já no método de formigueiro, as sementes caem conforme as vagens amadurecem, e consequentemente as suas sementes são liberadas no ponto de maturação fisiológica. Este tipo de coleta acaba por ser menos oneroso e demonstrou ser positivo em relação à qualidade das sementes, com porcentagens de germinação variando de 60,67 a 74,17% (Tabela 1). Matheus, Lopes e Corrêa (2011) também relatam que a coleta das sementes de Erythrina variegata deve ser realizada após as mesmas atingirem o ponto de maturidade fisiológica, o que auxiliará na qualidade das sementes. De acordo com Borges e Borges (1979) um dos fatores que influencia na qualidade das sementes é a maturação, sendo que diferentes tipos de coleta nem sempre coincidem com o pleno vigor da semente e a coleta realizada antes ou depois do amadurecimento do fruto pode influenciar negativamente na germinação. Neste sentido se faz necessário a realização de estudos quanto ao tipo de tratamento e tempo de imersão da semente para cada espécie e até mesmo entre diferentes lotes e procedências de uma mesma espécie, devido a variações que ocorrem na composição e na espessura dos tegumentos, podendo ocasionar diferentes respostas na germinação (Freire, Ataíde, Rouws, 2016; Berger, Ranal, Santana, 2014).
Houve diferença entre os métodos de produção de sementes para o índice de velocidade de germinação (IVG), com os melhores resultados para sementes provenientes de formigueiro, com valor máximo de 2,06, frente ao IVG das sementes de APS, de 0,86 Não foi verificada influência entre as concentrações de giberelinas testadas para o IVG (Gráfico 1). Diferentemente do observado no presente trabalho com sementes de Annona cherimola Mill. x A. squamosa L. foi verificado que o IVG foi significativamente superior nas concentrações mais elevadas de GA (750 e 1000 mg L-1) (Oliveira et al., 2010). Tanto o IVG quanto a germinação são ótimos parâmetros que denotam a qualidade das sementes, sendo verificado em sementes de Piptadenia viridiflora que o IVG foi o índice que melhor caracterizou a maturidade fisiológica das sementes (Pessoa et al., 2010).
Gráfico 1 - Índice de velocidade de germinação de sementes de Acacia mearnsii
provenientes de área de produção de sementes e formigueiro, submetidas à diferentes
concentrações de giberelina.
Com o método de produção de sementes em formigueiro foram observados maiores comprimentos de raízes (27,50 - 37,82 mm) quando comparado a APS (21,90 – 27,27mm), embora não tenha sido verificada diferença estatística com relação aos tratamentos com GA (Tabela 2). Em raiz de Arabidopsis foi constatado que após a proteína DELLA ter sido expressa, pela ação do gene gai o crescimento da raiz foi retardado em células endodérmicas de raiz, demonstrando que a endoderme representa o tecido primário que regula o crescimento dos órgãos e que a expansão das células endodérmicas limita a taxa de alongamento da raiz como um todo (Ubeda-Tomás et al., 2008). Mesmo que haja conhecimento de que a proteína DELLA exerça um bloqueio na resposta ao GA, ainda é pouco compreendido o papel da GA no desenvolvimento das raízes (Gou et al. 2010).
Os melhores resultados para o comprimento da parte aérea (15,34 e 16,01 mm) foram observados com a maior concentração de GA testada (750 mg L-1) (Tabela 3). Vários autores já relataram o efeito fisiológico do GA no crescimento da parte aérea (Lima et al., 2009; Peixoto et al., 2011). Além de relatos de que a giberelina acelera a germinação ela também é citada por atuar na modificação do crescimento e desenvolvimento de plantas, por funcionar na regulação da divisão e do alongamento das células de várias espécies e também por intermediar os efeitos de estímulos ambientais no desenvolvimento de plantas (Lima et al., 2009). O efeito da giberelina baseia-se na ativação de enzimas hidrolíticas de reservas nutritivas disponibilizando para o embrião compostos necessários para o seu crescimento e desenvolvimento. As giberelinas promovem a divisão e o alongamento celular ao promover o aumento da extensibilidade e da plasticidade da parede celular, atuando preferencialmente em células jovens e meristemáticas, culminando em uma melhor emergência das plântulas (Salisbury e Ross, 1991, 682 p.).

Os maiores comprimentos do hipocótilo ocorreram com o tratamento de 750 mg L-1 (35,57 e 35,74 mm) (Tabela 4). Bidadi et al. (2010) relatam que o ácido giberélico promove também a manutenção do estado fisiológico do hipocótilo e Peng e Harberd (2002) e Richards et al. (2001) sugerem que isso ocorre principalmente devido à ação do GA na síntese de enzimas hidrolíticas de reserva nutritiva, como a a amilase, produzindo efeitos no alongamento do hipocótilo. Podendo influenciar o crescimento de vários órgãos da planta, devido ao efeito relacionado à promoção do alongamento e divisão celular (Ferrari et al., 2007), podendo exercer influência tanto no hipocótilo, quanto na parte aérea.
De acordo com os resultados obtidos no presente trabalho com Acacia mearnsii conclui-se que a germinação, índice de velocidade de germinação (IVG) e comprimento da raiz não são afetados com as concentrações de giberelina testadas. A concentração de 750 mg L-1 de giberelina promove maiores comprimentos da parte aérea e hipocótilo. O método de produção de sementes em formigueiro apresenta melhor germinação, IVG e comprimento de raiz em relação ao método de produção em APS.
AGEFLOR – Associação Gaúcha de Empresas Florestais (2014). A indústria de Base Florestal no RS – Dados e fatos ano base 2014. Recuperado de http://www.ageflor.com.br/
Auer, C. G., Silva, R. (1992). Fixação de nitrogênio em espécies arbóreas. In: Cardoso, E. J. B. N., Tsi, M.; Neves, M. C. P. (eds.), Microbiologia do solo. (p. 160-167). Campinas: Universidade Estadual de São Paulo.
Berger, A. P. de A., Ranal, M. A., Santana, D. G. de (abril/junho, 2014) Variabilidade na dormência relativa dos diáporos de Lithraea molleoides (Vell.) Eng. Ciência Florestal, 24(2), 1-13.
Bidadi, H., Yamaguchi, S., Asahina M., Satoh S. (2010). Effects of shoot-applied gibberellin/gibberellin-biosynthesis inhibitors on root growth and expression of gibberellin biosynthesis genes in Arabidopsis thaliana. Plant Root, 4, 4-11.
Borges, E. E. L., Borges, C. G. (1979). Germinação de sementes de Copaifera langsdorffii Desf. provenientes de frutos com diferentes graus de maturação. Revista Brasileira de Sementes, 1(3), 45-47.
Dalastra, I. M., Pio, R., Entelmann, F. A., Scarpare Filho, J. A., Uliana, M. B., Werle, F. (maio/junho, 2010). Germinação de sementes de nogueira-macadâmia submetidas à incisão e imersão em ácido giberélico. Ciência e Agrotecnologia, 34(3), 641-645.
Dizarz, R., Corder, M. P. M. (2009). Multiplicação de gemas axilares de Acacia mearnsii De Wild. sob diferentes meios de cultura. Revista Árvore, 33(4), 599-606.
Engel, M. L., Higa, A. R., Alcantara, G. B. de, Flôres Junior, P. C., Soares, I. D. (2017). Enraizamento de miniestacas de diferentes clones de Acacia mearnsii De Wildeman com aplicação de AIB. Revista Espacios, 38(23), 8-19.
Ferrari, T. B., Ferreira, G., Boaro, C. S. F., Zucareli, V. (julho, 2007). Bioestimulante no crescimento de plântulas de maracujazeiro-doce. Revista Brasileira de Biociências, 5(2), 342-344.
Flores, I. F., Passamani, S., Bonow, R. N., Lemos, C. A. (2002). Tratamento de sementes com ácido giberélico e crescimento de plântulas de arroz (Oryza sativa, L). Revista da FZVA, 9(1), 73-78.
Fokuhl, G., Heinze, J., Poschlod, P. (2012). Myrmecochory by small ants – Beneficial effects through elaiosome nutrition and seed dispersal. Acta Oecologica, 38, 71-76.
Freire, J. M., Ataíde, D. H. dos S., Rouws, J. R. C. (2016). Superação de dormência de sementes de Albizia pedicellaris (DC.) L. Rico. Floresta e Ambiente, 23(2): 251-257.
Gou, J., Strauss, S. H., Tsai, C. J., Fang, K., Chen, Y., Jiang, X., Busov, V. B. (march, 2010). Gibberellins regulate lateral root formation in Populus through interactions with auxin and other hormones. The Plant Cell, 22(3), 623-639.
Handel, S. N., Beattie, A. J. (1990). Seed dispersal by ants. Science American, 263(1), 54-61.
IBÁ - Indústria Brasileira de Árvores (2016). Relatório anual 2016. Recuperado de http://iba.org/images/shared/Biblioteca/IBA_RelatorioAnual2016_.pdf
Lima, C. S. M., Betemps, D. L., Tomaz, Z. F. P., Galarça, S. P., Rufato, A. de R. (janeiro/dezembro, 2009). Germinação de sementes e crescimento de maracujá em diferentes concentrações do ácido giberélico, tempos de imersão e condições experimentais. Revista Brasileira de Agrociência,15,(1-4), 43-48.
Malavasi, M. M. (1988). Germinação de Sementes.. In: Piña Rodrigues, F. C. M. (eds.). Manual de Análise de Sementes Florestais. (p. 1-100). Campinas: Fundação Cargill.
Martins-Corder, M. P.; Borges Junior, N. (1999). Desinfestação e quebra de dormência de sementes de Acacia mearnsii de Wild. Ciência Florestal, 9(2), 1-7.
Matheus, M. T., Lopes, J. C., Corrêa, N. B. (outubro/dezembro, 2011). Maturação fisiológica de sementes de Erythrina variegata L. Ciência Florestal, 21(4), 619-627.
Mochiutti, S., Higa, A. R., Simon, A. A. (maio/ agosto, 2007). Susceptibilidade de ambientes campestres à invasão de acácia-negra (Acacia mearnsii de Wild.) no Rio Grande do Sul. Floresta, 37(2), 239-253.
Mora, A. L. (2002). Aumento da produção de sementes geneticamente melhoradas de Acacia mearnsii de Wild. (acácia-negra) no Rio Grande do Sul (Tese de doutorado). Programa de pós-graduação em Engenharia Florestal. Universidade Federal do Paraná. Curitiba.
Oliveira, M. C. de, Ferreira, G., Guimarães, V. F., Dias G. B. (junho, 2010). Germinação de sementes de atemoia (Annona Cherimola mill. x A. squamosa l.) cv ‘gefner’ submetidas a tratamentos com ácido giberélico (GA3) e ethephon1. Revista Brasileira de Fruticultura, 32(2), 544-553.
Peixoto, C. P., Sales, F. de J. S., Vieira, E. L., Passos, A. R., Santos, J. M. da S. dos, (2011). Ação da giberelina em sementes pré-embebidas de mamoneira. Comunicata Scientiae, 2(2), 70-75.
Peng, J., Harberd, N. P. (october, 2002). The role of GA-mediated signalling in the control of seed germination. Current Opinion in Plant Biology, 5(1), 376-381.
Pessoa, R. C., Matsumoto, S. M., Morais, O. M., Vale, R. S., Lima, J. M. (julho/agosto, 2010). Germinação e maturidade fisiológica de sementes de Piptadenia viridiflora (Kunth.) Benth relacionadas a estádios de Frutificação e conservação pós-colheita. Revista Árvore, 34(4), 617-625.
Pires, I. E., Resende, M. D. V. de, Silva, R. L., Resende Júnior, M. F. R. de (2011). Genética florestal. Viçosa: Arka.
RAS – Regras para análise de sementes (2009). Ministério da Agricultura Pecuária e Abastecimento. Recuperado de http://www.agricultura.gov.br/assuntos/insumos-agropecuarios/arquivos-publicacoes-insumos/2946_regras_analise__sementes.pdf
Richards, D. E., King, K. E., Ait-Ali, T., Harberd, N. P. (june, 2001). How gibberellin regulates plant growth and development: a molecular genetic analysis of gibberellin signaling. Annual review of plant physiology and Plant Molecular Biology, 52, 67-88.
Riegel, I., Moura, A. B. D., Morisso, F. D. P., Mello, F. de S. (maio/junho, 2008). Análise termogravimétrica da pirólise da acácia-negra (Acacia mearnsii de Wild.) cultivada no Rio Grande do Sul, Brasil, Revista Árvore , 32(3), 533 - 543.
Salisbury, F. B., Ross, C. W. (1991). Plant Physiology. Belmont: Wadsworth.
Scalon, S. de P. Q., Lima, A. A. de, Scalon Filho, H., Vieira, M. do C. (2009). Germinação de sementes e crescimento inicial de mudas de Campomanesia adamantium camb.: efeito da lavagem, temperatura e de bioestimulantes. Revista Brasileira de Sementes, 31(2), 96-103.
Silva, F. de A. S., Azevedo, C. A. V. de, (september, 2016). The Assistat software version 7.7 and its use in the analysis of experimental data. African Journal of Agricultural Research, 11(39), 3733-3740.
Stein, V. C., Paiva, R., Soares, F. P., Nogueira, R. C., Silva, L. C., Emrich, E. (novembro/dezembro, 2007). Germinação in vitro e ex vitro de Inga vera willd. subsp. affinis (DC.) T.D. Penn. Ciência e Agrotecnologia, 31(6), 1702-1708.
Ubeda-Tomás, S., Swarup, R., Coates, J., Swarup, K., Laplaze, L., Beemster, G. T. S., Hedden, P., Bhalerao, R., Bennett, M. J. (MAY, 2008). Root growth in Arabidopsis requires gibberellin/DELLA signalling in the endodermis. Nature Cell Biology, 10(5).
1. Aluno de graduação do Curso de Engenharia Florestal, Universidade Federal do Paraná, Curitiba – PR.
2. Doutorando do Curso de Pós Graduação em Engenharia Florestal, Universidade Federal do Paraná, Curitiba – PR.
3. Professor do Departamento de Ciências Florestais, Setor de Ciências Agrárias, Universidade Federal do Paraná, Curitiba – PR.
4. Professor do Departamento de Ciências Florestais, Setor de Ciências Agrárias, Universidade Federal do Paraná, Curitiba – PR.