![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 46) Ano 2017. Pág. 27
Graciliano Galdino Alves dos SANTOS 1 ; Mariana OLIVEIRA 2; Izildinha Souza MIRANDA 3; Danielle MITJA 4; Natália MAFRA 5; Anne-Elisabeth LAQUES 6
Recebido: 24/05/2017 • Aprovado: 22/06/2017
RESUMO: O presente estudo objetiva avaliar o efeito das métricas de uma paisagem antropizada em diferentes escalas sobre os diferentes estratos da vegetação. Foram realizadas RDA entre as variáveis da vegetação vs. métricas de paisagem. As métricas proporção de área não-florestada e densidade de fragmentos influenciaram os três estratos da vegetação em todas as escalas, refletindo o forte efeito da perda de hábitat e fragmentação. As métricas relacionadas à ambientes bem preservados tem efeito consistente na escala de 2000 m. |
ABSTRACT: The present study aims to compare the metrics effect in a anthropized landscape at different scales and forest layers. Redundancy analyses were made between plant variables vs. landscape metrics. The proportion of non-forested area and fragments density affected the three layers in all scales, reflecting the strong effect of habitat loss and fragmentation. The metrics related to well-preserved environments were consistent at 2000 m scale. |

Os padrões de distribuição espacial da biodiversidade, que refletem os gradientes ambientais, há décadas são de interesse da Ecologia (Whittaker, 1956). Porém, só em meados dos anos 80, com a necessidade de definir critérios quantitativos (métricas) para descrever as paisagens e com o desenvolvimento de técnicas de sensoriamento remoto e geoprocessamento, começaram a ser compreendidos os efeitos da composição e configuração da paisagem sobre os processos ecológicos (Turner e Garnder, 2015 ).
Em paisagens de florestas com intensa exploração humana, a substituição de áreas de vegetação nativa por diversos tipos de usos da terra, tende a isolar os remanescentes florestais afetando os processos como a dispersão de sementes e polinização (Prevedello e Vieira, 2010; Breed et al., 2015). Nessas paisagens também ocorre a redução da área dos fragmentos florestais, alterando as condições microclimáticas dos mesmos devido ao aumento da influência do efeito de borda (i.e., intensificação da entrada de luz, vento e fogo que causam aumento de temperatura e perda de umidade) (Murcia, 1995; Camargo e Kapos, 1995; Zuidema et al., 1996). Essas novas condições reduzem a taxa de recrutamento de plântulas tolerantes à sombra (Benitez-Malvido, 1998), reduzem o fitness de árvores devido ao estresse fisiológico (Alvarez-Buylla et al., 1996) e aumentam a mortalidade de grandes árvores (Laurance et al., 2000; Oliveira et al., 2008).
As métricas de paisagem têm sido amplamente usadas para compreender como os diferentes processos ecológicos são afetados pela fragmentação das florestas, ex. o sucesso na dispersão (Gustafson e Gardner, 1996; Tischendorf, 2001), os padrões de abundância (Mc Garigal; Mc Comb, 1995) e probabilidade de sobrevivência (Fahrig, 1998).
As métricas da paisagem e a escala espacial devem ser cuidadosamente escolhidas nesses estudos (Wu et al., 2002), já que a subjetividade desses procedimentos pode levar a interpretações errôneas dos reais padrões e processos ecológicos de uma área. Schindler et al., (2008) identificaram 4 métricas de paisagem (de 55 analisadas) que tiveram influência sobre a cobertura da terra em diferentes escalas (250 ha e 1000 ha), para definir quais seriam apropriadas para um plano de monitoramento no Dadia National Park, Grécia. Já Fan e Mynt (2014) tiveram sucesso em relacionar a vegetação com somente 06 métricas em diferentes escalas.
Porém, importantes padrões de comportamento da biodiversidade em escalas maiores, como o das espécies dominantes e hiperdominantes (Ter Steege et al., 2013) permanecem sem maiores evidências de como são afetados pelos novos arranjos espaciais causados pela influência humana. O presente estudo objetiva avaliar o efeito da escala e configuração de uma paisagem antropizada sobre os diferentes estratos da vegetação, uma vez que eles representam grupos de espécies que são influenciados por diferentes fatores na escala local (Puig, 2008).
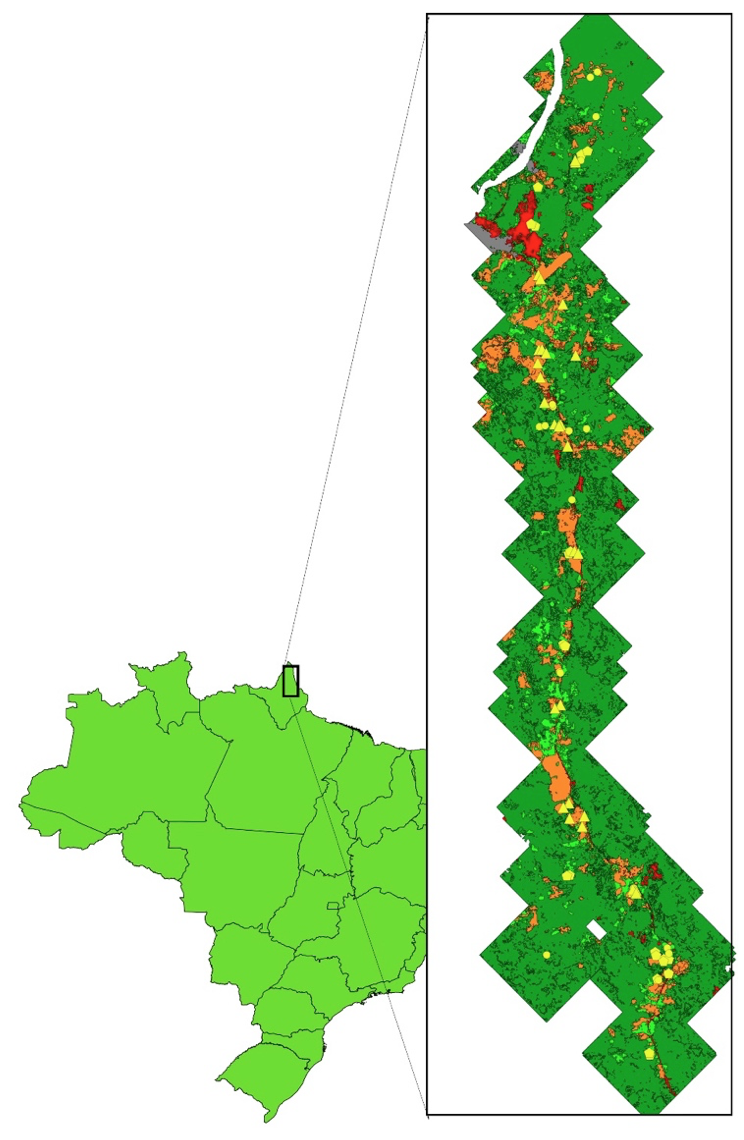
O estudo foi realizado no nordeste da Amazônia, no município de Oiapoque, estado do Amapá, limite do Brasil com Guiana Francesa. A temperatura média é 26,5ºC e a precipitação média anual é 2500 mm, com estação seca entre setembro e novembro (Souza e Cunha, 2010). Os solos predominantes na área são o latossolo amarelo, aluminizados, areno-argilosos, arenosos, argilo-siltosos e conglomerados, cobertos predominantemente pela floresta densa de terras baixas (IBGE, 2004).
As parcelas foram implantadas em propriedades particulares ao longo da BR-156, após a seleção baseada em entrevistas com proprietários e visita às áreas, visando abranger as coberturas vegetais mais comuns da região (Figura 1), que é dominada por florestas primárias (59,6% da área do município), mas também apresenta florestas secundárias (12,6%), pastagens (5,6%) e roças (1,5%).
Foram utilizadas imagens Landsat 8 OLI/TIRS Surface Reflectance (órbitas-ponto 226/057 e 226/058) do ano de 2013, quando também foi realizado o inventário da vegetação. As imagens foram classificadas em floresta, capoeira, pasto, solo exposto, água, nuvem e sombra. A classificação foi feita a partir das bandas 6-5-4 utilizando 70 pontos do inventário da da vegetação, através do classificador K-Means do software Spring 5.4.1.
Para contornar o efeito de cobertura de nuvens, primeiro foram classificadas 4 imagens de cada órbita-ponto; em seguida, foram retiradas as categorias nuvens e sombra das classificações; e por fim, as classificações foram unificadas gerando uma única imagem classificada, sem nuvens, para cada órbita-ponto.
A classificação foi validada utilizando imagens de alta resolução (2,5 m x 2,5 m) do radar SAR.
Figura 1. Localização da área de estudo no Estado do Amapá. Os pontos
em amarelo representam as parcelas inventariadas.
As métricas da paisagem foram calculadas através do software Fragstats 4.2.1. Foi utilizado o método de amostragem por pontos (= parcelas dos inventários realizados em campo); os mesmos serviram como centro para os quadrados de diferentes escalas, nos quais as métricas foram calculadas.
Foram selecionadas 18 métricas que refletem as diferentes características da paisagem (Tabela 1) conforme recomendado por Mc Garigal (2017), acrescentada a proporção de área não florestada (prop_nfor), que é a soma da área das coberturas não-florestadas dividido pela área total. As métricas foram calculadas em áreas quadrangulares com seis diferentes escalas: 200 m, 500 m, 800 m, 1000 m, 1500 m, 2000 m de aresta do quadrado. A menor escala (200 m) foi baseada na distância máxima de dispersão de sementes anemocóricas das espécies tropicais, que é aproximadamente 194 m (Augspurger, 1986).
Tabela 1. Métricas da paisagem utilizadas de acordo com o grupo a que pertencem.
Grupo |
Métrica |
Descrição |
Área-Borda |
prop_nfor |
Proporção de área não-florestada |
Área-Borda |
area_md |
Área dos Fragmentos (mediana) |
Área-Borda |
gyrate_md |
Raio de Rotação (mediana) |
Área-Borda |
lpi |
Índice de Maior Fragmento |
Dispersão - Justaposição |
cohesion |
Índice de Coesão dos Fragmentos |
Dispersão - Justaposição |
contag |
Contágio |
Diversidade |
shdi |
Índice de Diversidade Shannon |
Diversidade |
sidi |
Índice de Diversidade Simpson |
Diversidade |
shei |
Índice de Equabilidade de Shannon |
Forma |
pafrac |
Índice de Dimensão Fractal por Área e Perímetro |
Forma |
circle_md |
Círculo Circunscrito Relacionado (mediana) |
Isolamento |
enn_md |
Distancia Euclidiana do Vizinho Mais Próximo (mediana) |
Isolamento |
prox_md |
Índice de Proximidade (mediana) |
Isolamento |
connect |
Índice de conectividade |
Núcleo |
cai_md |
Índice de Áreas Núcleo |
Núcleo |
dcore_md |
Distribuição de Áreas Núcleo Disjuntas (mediana) |
Subdivisão |
pd |
Densidade de Fragmentos |
Subdivisão |
division |
Índice de Divisão da Paisagem |
Subdivisão |
mesh |
Tamanho Efetivo da Malha |
Foram amostradas 70 parcelas de 10 m x 50 m em áreas de 16 pequenos produtores familiares; 20 parcelas em florestas primárias, 20 em florestas secundárias e 30 em pastagens. Em toda a extensão da parcela foram inventariados todos os indivíduos com DAP > 10 cm (Estrato Superior); e dentro destas parcelas foram alocadas sub-parcelas de 5 x 50 m para inventariar os indivíduos com DAP < 10 cm e altura > 2.0 m (Estrato médio); por fim, um transecto de 1 x 50 m foi colocado no centro da parcela para inventariar os indivíduos com 0.10 m < altura < 2.0 m (Estrato inferior).
Os indivíduos foram identificados por um parataxônomo ou por comparação com exsicatas depositadas no Herbário João Murça Pires do Museu Paraense Emílio Goeldi (MPEG). A correção dos nomes botânicos foi realizada de acordo com a plataforma The Plant List (www.theplantlist.org). As espécies foram classificadas em pioneiras ou tolerantes à sombra conforme Do Vale et al. (2015).
Foram consideradas espécies dominantes as mais abundantes, que detiveram 50% dos indivíduos da parcela. Foram utilizadas 21 variáveis que representam a estrutura da vegetação (Tabela 2).
Para verificar qual efeito da configuração das paisagens sobre a vegetação, foram realizadas dezoito análises de redundância (RDA) entre a matriz de vegetação e a matriz de métricas da paisagem, uma para cada escala (200 m, 500 m, 800 m, 1000 m, 1500 m e 2000 m) e para cada estrato da vegetação (superior, médio e inferior). Para as análises, foi aplicada a transformação Z ((X – Média) / Desv.Padrão) em todas as variáveis da matriz de vegetação. Em seguida, foram realizados testes de permutação (105 permutações, α = 5%) similares à ANOVA para identificar quais métricas tiveram efeito significativo sobre as ordenações. As análises foram realizadas no R 3.3.2. (R Foundation for Statistical Computing, Vienna, AT) utilizando o pacote vegan.
Tabela 2. Variáveis da vegetação utilizadas para a Análise de Redundância
Variável |
Descrição |
abu_inf |
abundância do estrato inferior |
riq_inf |
riqueza do estrato inferior |
dom_inf |
número de espécies dominantes do estrato inferior |
pdom_inf |
proporção de espécies dominantes do estrato inferior |
J_inf |
equibilidade do estrato inferior |
pdomT _inf |
proporção de espécies tolerantes dominantes do estrato inferior |
pdomP_inf |
proporção de espécies pioneiras dominantes do estrato inferior |
abu_med |
abundância do estrato médio |
riq_med |
riqueza do estrato médio |
dom_med |
número de espécies dominantes do estrato médio |
pdom_med |
proporção de espécies dominantes do estrato médio |
J_med |
equibilidade do estrato médio |
pdomT_med |
proporção de espécies tolerantes dominantes do estrato médio |
pdomP_med |
proporção de espécies pioneiras dominantes do estrato médio |
abu_sup |
abundância do estrato superior |
riq_sup |
riqueza do estrato superior |
dom_sup |
número de espécies dominantes do estrato superior |
pdom_sup |
proporção de espécies dominantes do estrato superior |
J_sup |
equibilidade do estrato superior |
pdomT_sup |
proporção de espécies tolerantes dominantes do estrato superior |
pdomP_sup |
proporção de espécies pioneiras dominantes do estrato superior |
Todas as análises de redundância mostraram que os tipos de cobertura formam grupos distintos (Figura 2-4), tendo o percentual de explicação das métricas sobre a variância dos dados de vegetação entre 37,4% e 51,1%. Esses percentuais, que refletem o ajuste do modelo (R2), são comuns para modelos que avaliam os padrões de distribuição espacial da biodiversidade. Dormann (2007), avaliando 20 artigos sobre padrões de distribuição espacial de diferentes grupos ecológicos, encontrou modelos estatísticos não-espaciais (como o utilizado no presente trabalho) com R2 médio de 0,43% e para modelos espaciais, média de R2= 0,49%.
Nos três estratos (Fig. 2-4), quase todas as variáveis biológicas tiveram maiores valores associados às florestas conservadas. Porém, a proporção de dominantes pioneiras apresentou maiores valores associados aos pastos (pdomP_inf) e às florestas secundárias no estrato médio (pdomP_med) e superior (pdomP_sup). Esse padrão indica que o aumento da dominância de espécies está associado a ambientes mais degradados e que, no estrato médio e superior (a partir dos 500 m), a dominância da comunidade está correlacionada à dominância das espécies pioneiras. O aumento da dominância de pioneiras é bem documentado em paisagens antropizadas, pois a perturbação das áreas altera as condições ambientais favorecendo o estabelecimento de espécies pioneiras (Santo-Silva et al., 2016).
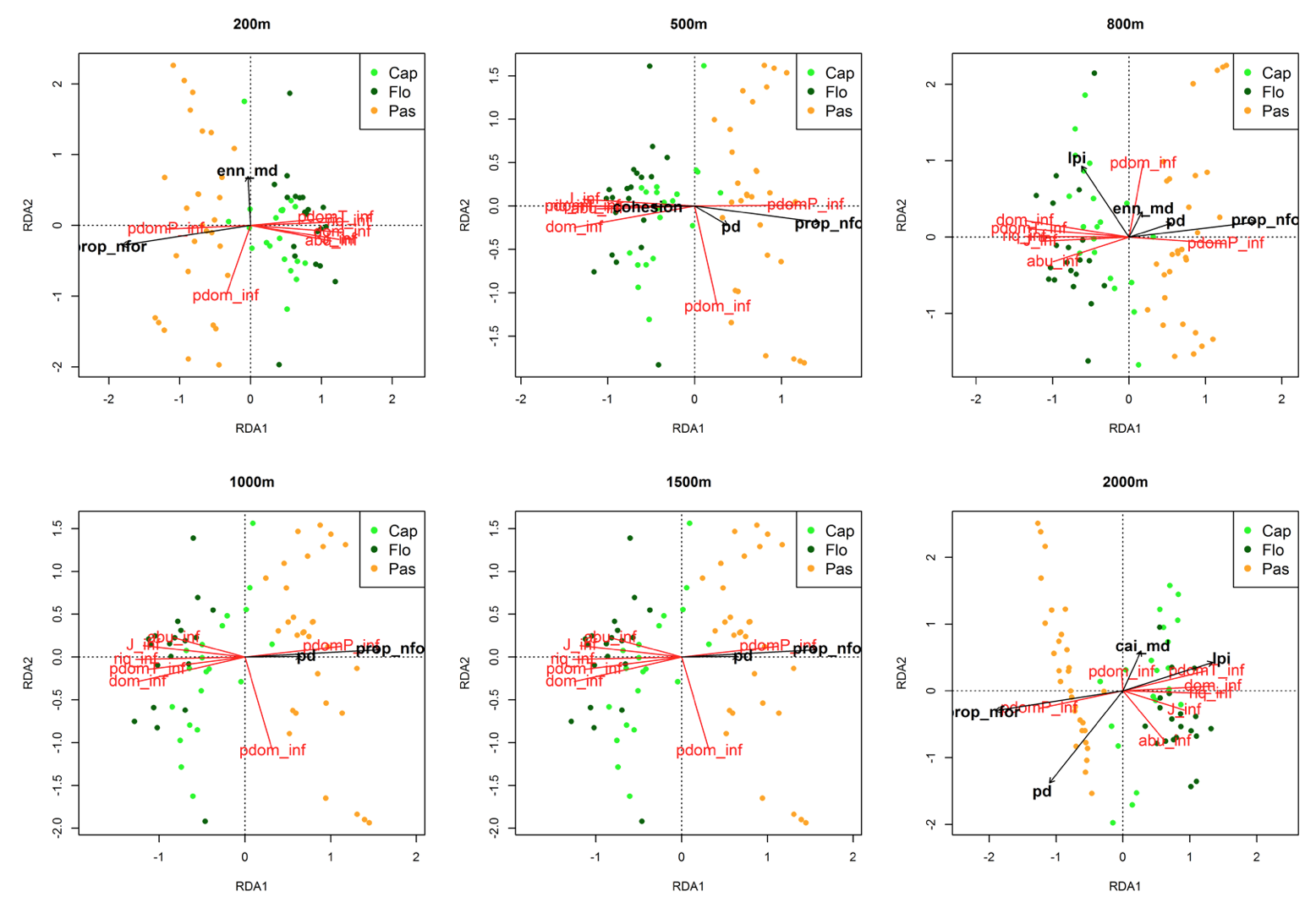
Entre as 19 métricas analisadas foram encontradas oito métricas significativas nas análises de redundância (Tabela 3). A proporção de áreas não florestadas (prop_nfor) e a densidade de fragmentos (pd; a partir de 500 m) foram correlacionadas positivamente à proporção de espécies pioneiras dominantes do estrato inferior (pdomP_inf; Fig. 2), e foram correlacionadas negativamente à maioria das outras variáveis da vegetação (Fig. 2-4).
Esse padrão da importância da quantidade de área sobre a biodiversidade, independente da configuração da paisagem, também foi encontrado por Quesnele et al. (2013) ao avaliarem o efeito da paisagem sobre aves e tartarugas. O resultado também corrobora, parcialmente, com a hipótese da quantidade de hábitat (Fahrig, 2013), i.e. somente a quantidade de habitat, independente do grau de isolamento e do tamanho dos fragmentos, tem influência sobre a riqueza de espécies. Porém, ao contrário do proposto por Fahrig (2013), a densidade de fragmentos (pd) também teve forte influência sobre a vegetação.
Figura 2. RDA da matriz da vegetação vs. métricas com efeito significativo no estrato inferior.
As siglas das métricas da paisagem e das variáveis da vegetação estão conforme as tabelas 1 e 2.
-----
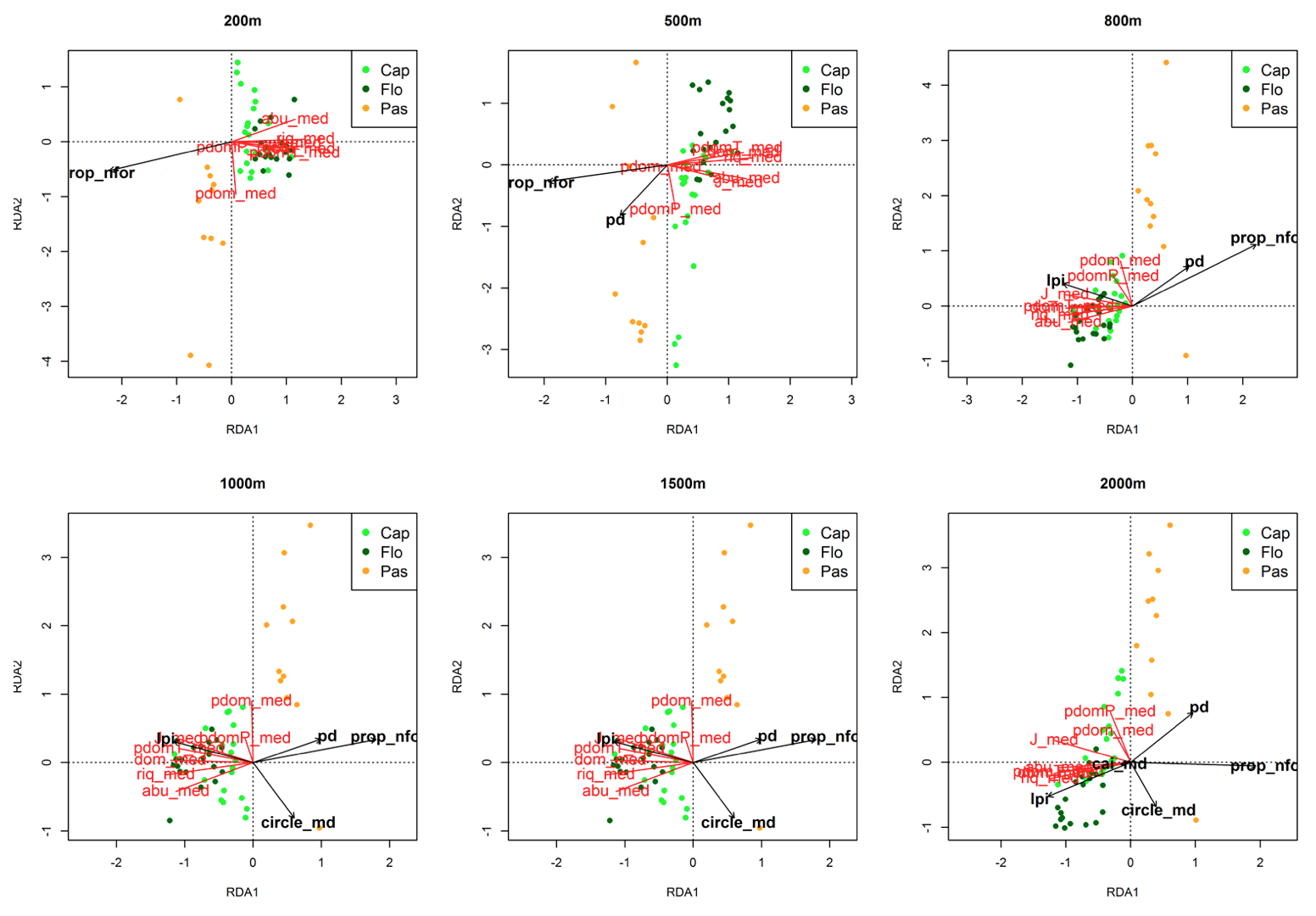
Figura 3. RDA da matriz da vegetação vs. métricas com efeito significativo no estrato médio.
As siglas das métricas da paisagem e das variáveis da vegetação estão conforme as tabelas 1 e 2.
-----
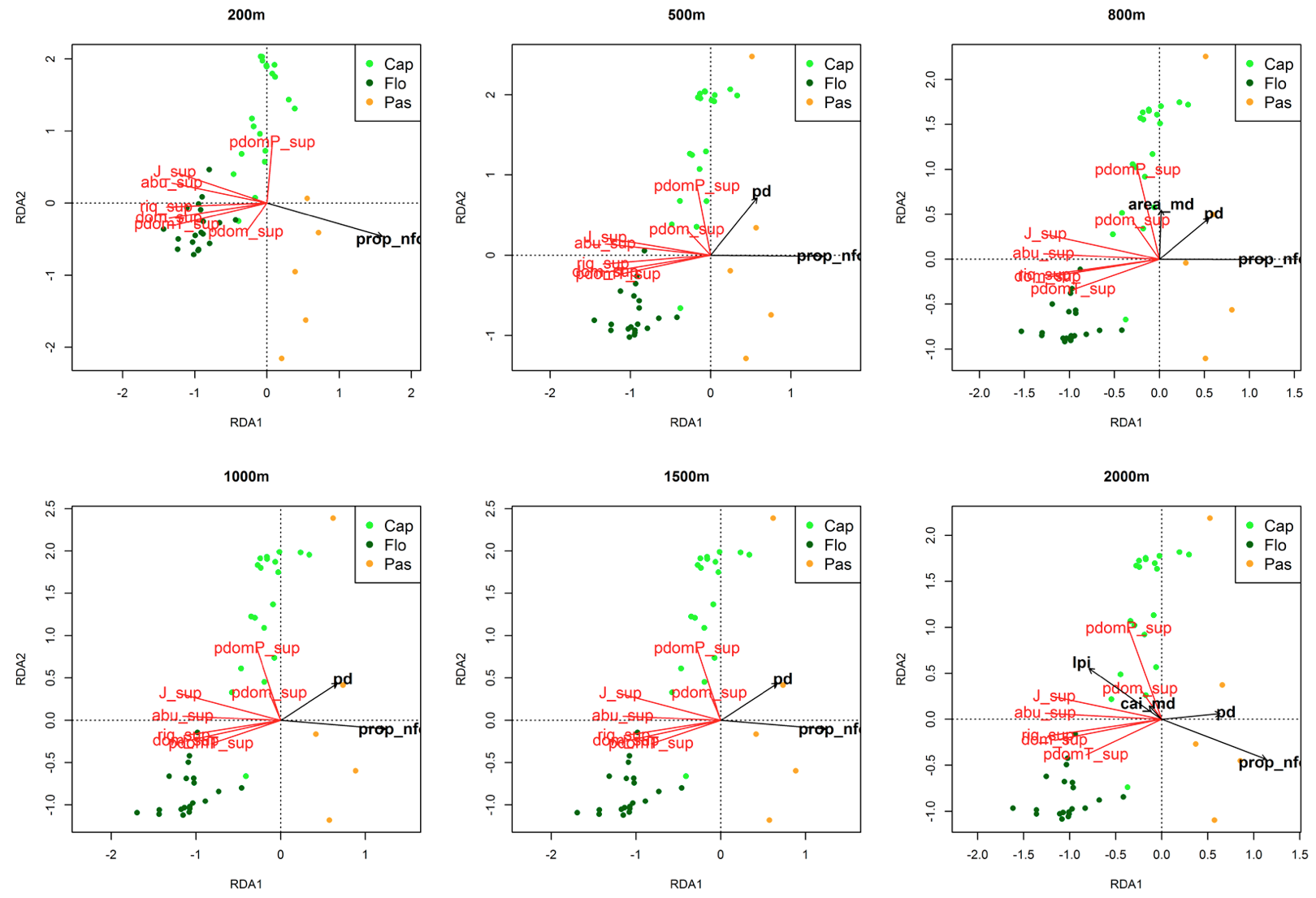
Figura 4. RDA da matriz da vegetação vs. métricas com efeito significativo no estrato superior.
As siglas das métricas da paisagem e das variáveis da vegetação estão conforme as tabelas 1 e 2.
-----
Tabela 3. Métricas da paisagem com efeito significativo nas RDA em
cada estrato (linhas) por escala da paisagem (em metros).
Escala |
Estrato da Vegetação |
||
Inferior |
Médio |
Superior |
|
200 |
prop_nfor enn_med |
prop_nfor |
prop_nfor |
500 |
prop_nfor pd cohesion |
prop_nfor pd |
prop_nfor pd |
800 |
prop_nfor pd Ipi enn_med |
prop_nfor pd lpi |
prop_nfor pd
area_md |
1000 e 1500 |
prop_nfor pd |
prop_nfor pd circle_md lpi |
prop_nfor pd |
2000 |
prop_nfor pd lpi cai_md |
prop_nfor pd lpi cai_md circle_md |
prop_nfor pd lpi cai_md |
O efeito significativo da proporção de áreas não florestadas (prop_nfor) e a densidade de fragmentos (pd) em todas as análises de redundância indica como a perda de área florestada e fragmentação dos hábitats, independente da escala, tem forte efeito sobre a vegetação. Esse resultado reforça como a perda de habitat e fragmentação são fatores preponderantes para a perda de espécies (Rybicki e Hanski , 2013; Barlow et al., 2016 ).
No estrato inferior, a correlação positiva entre proporção de espécies pioneiras dominantes (pdomP) com proporção de áreas não florestadas (prop_nfor) e a densidade de fragmentos (pd) e a relação negativa com as demais variáveis biológicas, refletem como as alterações na paisagem afetam diretamente os processos que influenciam na dispersão e recrutamento de ervas e plântulas de árvores. Santo-Silva et al. (2013) identificaram efeito significativo do tamanho dos fragmentos sobre o estrato herbáceo em remanescentes de Floresta Atlântica, com pequenos fragmentos dominados por poucas espécies pioneiras. Na região do Oiapoque, as áreas com fragmentos de menor porte são também aquelas com alta densidade de fragmentos (pd).
O índice de maior fragmento (lpi), i.e. percentual de área ocupada pelo maior fragmento, na escala de 2000 m, foi correlacionado positivamente com a proporção de espécies dominantes tolerantes (pdomT) no estrato inferior e médio, e com a proporção de espécies dominantes (pdom) e com a proporção de espécies dominantes pioneiras (pdomP) no estrato superior. O índice de área núcleo (cai_md), teve padrão similar ao índice de maior fragmento, porém só teve efeito significativo no estrato inferior e superior. No estrato médio, o Círculo Circunscrito Relacionado (que mede quão circular é a forma do fragmento) foi correlacionada negativamente com a proporção de espécies dominantes.
O fato do índice de maior fragmento e do índice de área núcleo estarem associados à dominância de pioneiras no estrato superior sugere que, na região do Oiapoque, áreas com grandes fragmentos sofrem pressões antrópicas há muito tempo, pois a atual presença das árvores adultas no estrato superior é reflexo dos fatores ambientais e bióticos de períodos mais antigos, que atuaram no momento do estabelecimento delas (Gonzaga et al., 2013). Porém, a alta correlação positiva de dessas mesmas métricas da paisagem com a dominância das espécies tolerantes (pdomT) nos estratos inferior e médio, mostra que grandes fragmentos mantêm a diversidade das espécies tolerantes em seus estágios iniciais de crescimento.
A correlação negativa de Círculo Circunscrito Relacionado (circle_md) com a dominância de espécies no estrato médio pode refletir o impacto das mudanças microclimáticas causadas pelo efeito das bordas dos fragmentos. Como ilustrado por Gascon et al. (2000), a regeneração das árvores (que se encontra no estrato inferior e médio) em florestas fragmentadas é afetada pela intensidade do efeito de borda. Logo, nas áreas com fragmentos menos circulares, teremos efeito de borda mais acentuado e maior proporção de espécies dominantes, provavelmente em sua maioria pioneiras.
As demais métricas, como o índice de Coesão dos Fragmentos (cohesion) e área dos fragmentos (area_md), só apareceram em uma escala, e não tiveram efeitos claramente relacionados às variáveis biológicas.
Quando analisadas as ordenações em diferentes escalas, não foi observado um gradiente de influência de diferentes métricas à medida que a escala aumenta. Em todos os estratos e escalas, a proporção de áreas não florestadas (prop_nfor) teve efeito significativo nas análises de redundância (Tabela 3). Outra variável que teve efeito significativo em todos os estratos, a partir de 500 m, foi a densidade de fragmentos (pd).
As diferentes respostas das métricas da paisagem às diferentes escalas são bem descritas por Wu et al. (2002), que identificou três tipos de resposta: as previsíveis, com influência clara da escala, facilmente descritas por modelos matemáticos simples (proporção de áreas não florestadas e densidade de fragmentos se encaixam nesse grupo); as tipo-escada, que em certos intervalos deixam de responder ao aumento da escala; e as erráticas, sem padrão claro de resposta. Ao aumentarmos a escala, também aumentamos a complexidade da configuração da paisagem e quais processos ecológicos podem ser explicados pelos arranjos espaciais (Tscharntke et al., 2005).
No estrato inferior, o número de métricas aumenta gradualmente até 800 m, mas em 1000 m e 1500 m somente proporção de áreas não florestadas e densidade de fragmentos influenciaram a ordenação. Provavelmente, alguns processos ecológicos mais influentes sobre as comunidades do estrato inferior, como a dispersão de sementes (Nathan e Muller-Landau, 2000), afetam de forma mais significativa até 800 m. A distância euclidiana do vizinho mais próximo (enn_md) e a medida de conectividade entre fragmentos (cohesion), por exemplo, são as métricas que tem efeito significativo até os 800 m, indicando que o efeito da distância e conectividade entre fragmentos, que influenciam diretamente na dispersão de sementes, sejam mais forte até essa distância.
No estrato médio, a proporção de áreas não florestadas (prop_nfor) e densidade de fragmentos (pd) foram significativas até 500 m, depois foi acrescentado a área do maior fragmento na paisagem (lpi) aos 800 m, Círculo Circunscrito Relacionado (circle_md) a partir dos 1000 m e o índice de Áreas Núcleo (cai_md) aos 2000 m. No estrato superior, a proporção de áreas não florestadas (prop_nfor) e densidade de fragmentos (pd) foram significativas até 1500 m e na escala de 2000 m foi observado o efeito de área do maior fragmento na paisagem (lpi) e índice de Áreas Núcleo (cai_md). A ausência de outras métricas além da proporção de áreas não florestadas e a densidade de fragmentos influenciando a vegetação do estrato superior até a escala de 2000 m indica que as métricas só vão influenciar as características das comunidades de árvores adultas, e podem ser úteis na compreensão de algum processo ecológico, em escalas mais amplas.
O efeito da proporção de área do maior fragmento na paisagem (Ipi) e do percentual de áreas núcleo na paisagem (cai_md) em todos os estratos na escala de 2000 m reflete que as métricas ligadas à boa qualidade do ambiente parecem ter efeito consistente sobre toda a comunidade vegetal em maiores escalas. Esse resultado reforça a necessidade de planejamentos de conservação em grandes escalas, utilizando unidades de paisagem com no mínimo 4 km2 (2000 m x 2000 m) para garantir paisagens com características que contribuam para manutenção da biodiversidade e processos ecológicos.
Duas métricas, a proporção de áreas não florestadas (prop_nfor) e densidade de fragmentos (pd), tiveram forte influencia sobre a vegetação, mostrando como a perda de habitat e a fragmentação afetam todos os estratos das comunidades vegetais. A maioria das variáveis ligadas à diversidade da vegetação tiveram maiores valores associados às florestas conservadas. A proporção de espécies dominantes pioneiras apresentou maiores valores associados aos pastos (pdomP_inf) e às florestas secundárias no estrato médio (pdomP_med) e superior (pdomP_sup).
Cada estrato foi influenciado por diferentes métricas em diferentes escalas: métricas ligadas ao processo de dispersão têm influencia sobre o estrato inferior até 800 m, o estrato médio é afetado pelo tamanho e forma dos fragmentos da paisagem a partir dos 800 m e as árvores adultas (estrato superior) só parecem ser influenciadas pela configuração da paisagem a partir dos 2000m. Esse resultado indica que, para estudos de comunidades vegetais em paisagen antropizadas é necessário considerar as unidades de paisagem de tamanho mínimo de 4 km2.
Este estudo foi financiado pela Superintendência do Desenvolvimento da Amazônia SUDAM. O Conselho Nacional de Ciência e Tecnologia (CNPq) concedeu a bolsa de estudo a Graciliano Galdino Alves dos Santos (processo 140880/2014-3). Nós agradecemos a Antonio Carlos “Beleza”, Salustiano Vilar Neto, Eduardo Leal, Elayne Braga, Tâmara Lima pela ajuda no trabalho de campo.
Alvarez-Buylla , E. R., Garcia-Barrios, R., Lara-Moreno, C., & Martinez-Ramos, M. (1996). Demographic and Genetic Models in Conservation Biology: Applications and Perspectives for Tropical Rain Forest Tree Species. Annual Review of Ecology and Systematics, 27(1), 387-421.
Augspurger, C. K. (1986). Morphology and dispersal potential of wind-dispersed diaspores of neotropical trees . American Journal of Botany, 353-363.
Barlow, J., Lennox, G. D., Ferreira, J., Berenguer, E., Lees, A. C., Mac Nally, R., ... Parry, L. (2016). Anthropogenic disturbance in tropical forests can double biodiversity loss from deforestation. Nature, 535(7610), 144-147.
Benitez‐Malvido, J. (1998). Impact of forest fragmentation on seedling abundance in a tropical rain forest. Conservation Biology, 12(2), 380-389.
Breed, M. F., Ottewell, K. M., Gardner, M. G., Marklund, M. H. K., Dormontt, E. E., & Lowe, A. J. (2015). Mating patterns and pollinator mobility are critical traits in forest fragmentation genetics . Heredity, 115(2), 108–114.
Camargo, J. L. C. & Kapos, V. (1995). Complex edge effects on soil moisture and microclimate in central Amazonian Forest . Journal of Tropical Ecology, 11(2), 205-221.
Do Vale, I., Miranda, I. S., Mitja, D., Grimaldi, M., Nelson, B. W., Desjardins, T., & Costa, L. G. S. (2015). Tree Regeneration Under Different Land-Use Mosaics in the Brazilian Amazon’s “Arc of Deforestation”. Environmental management, 56(2), 342-354.
Dormann, C. F. (2007). Effects of incorporating spatial autocorrelation into the analysis of species distribution data. Global Ecology and Biogeography, 16(2), 129-138.
Fahrig, L. (2013). Rethinking patch size and isolation effects: the habitat amount hypothesis. Journal of Biogeography, 40(9), 1649-1663.
Fahrig, L. (1998). When does fragmentation of breeding habitat affect population survival? Ecological Modelling, 105(2), 273-292.
Fan, C. & Myint, S. (2014). A comparison of spatial autocorrelation indices and landscape metrics in measuring urban landscape fragmentation . Landscape and Urban Planning, 121, 117-128.
Gascon, C., Williamson, B. & Fonseca, G.A.B. (2000). Receding forest edges and vanishing reserves . Science, 288(5470), 1356-1358.
Gonzaga, A. P. D., Pinto, J. R. R., Machado, E. L. M. & Felfili, J. M. (2013). Similaridade florística entre estratos da vegetação em quatro Florestas Estacionais Deciduais na bacia do Rio São Francisco. Rodriguésia, 64(1), 11-19.
Gustafson, E. J. & Gardner, R. H. (1996). The effect of landscape heterogeneity on the probability of patch colonization . Ecology 77(1), 94-107.
IBGE - Instituto Brasileiro de Geografia e Estatística (2004). Mapa de Vegetação do Brasil, 2ª edição. Rio de Janeiro, Brazil: Governo Federal do Brasil.
Laurance, W. F., Delamonica, P., Laurance, S. G., Vasconcelos, H. L. & Lovejoy, T. E. (2000). Conservation: rainforest fragmentation kills big trees . Nature, 404(6780) 836.
McGarigal, K., (2017). Landscape Metrics. Recuperado de http://www.umass.edu/landeco/teaching/landscape_ecology/schedule/chapter9_metrics.pdf
McGarigal, K. & McComb, W. C. (1995). Relationships between landscape structure and breeding birds in the Oregon Coast Range . Ecological monographs, 65(3), 235-260.
Murcia, C. (1995). Edge effects in fragmented forest: implications for conservation. Trends in Ecology and Evolution, 10(2), 58-62.
Nathan, R., & Muller-Landau, H. C. (2000). Spatial patterns of seed dispersal, their determinants and consequences for recruitment. Trends in ecology & evolution, 15(7), 278-285.
Oliveira, M. A., Santos, A. M. M., & Tabarelli, M. (2008). Profound impoverishment of the large-tree stand in a hyper-fragmented landscape of the Atlantic forest. Forest Ecology and Management, 256(11), 1910-1917.
Prevedello, J. A. & Vieira, M. V. (2010). Does the type of matrix matter? A quantitative review of the evidence. Biodiversity Conservation, 19(5), 1205–1223.
Puig, H. (2008). A floresta tropical úmida. São Paulo :Editora UNESP.
Quesnelle, P. E., Fahrig, L., & Lindsay, K. E. (2013). Effects of habitat loss, habitat configuration and matrix composition on declining wetland species. Biological Conservation, 160, 200-208.
Rybicki, J., & Hanski, I. (2013). Species–area relationships and extinctions caused by habitat loss and fragmentation. Ecology letters, 16(s1), 27-38.
Santo-Silva , E. E., Almeida, W. R., Melo, F. P., Zickel, C. S., & Tabarelli, M. (2013). The nature of seedling assemblages in a fragmented tropical landscape: implications for forest regeneration. Biotropica, 45(3), 386-394.
Santo-Silva, E. E., Almeida, W. R., Tabarelli, M., & Peres, C. A. (2016). Habitat fragmentation and the future structure of tree assemblages in a fragmented Atlantic forest landscape. Plant Ecology, 217(9), 1129-1140.
Schindler, S., Poirazidis, K. & Wrbka, T. (2008). Towards a core set of landscape metrics for biodiversity assessments: a case study from Dadia National Park, Greece. Ecological indicators. 8(5), 502-514.
Souza, E. B. & Cunha, A. C. (2010). Climatologia de Precipitação no Amapá e Mecanismos Climáticos de Grande Escala. En A. C. Cunha, E. B. Souza & H. F. A. Cunha (eds.). Tempo, clima e recursos hídricos: resultados do Projeto REMETAP no Estado do Amapa. Macapá, Brazil:IEPA.
Ter Steege, H., Pitman, N. C., Sabatier, D., Baraloto, C., Salomão, R. P., Guevara, J. E., ... Monteagudo, A. (2013). Hyperdominance in the Amazonian tree flora. Science, 342(6156), 1243092.
Tischendorf, L. (2001). Can landscape indices predict ecological processes consistently? Landscape ecology, 16(3), 235-254.
Tscharntke, T., Klein, A. M., Kruess, A., Steffan‐Dewenter, I., & Thies, C. (2005). Landscape perspectives on agricultural intensification and biodiversity–ecosystem service management. Ecology letters, 8(8), 857-874.
Turner, M. G. & Gardner, R. H. (2015). Landscape Ecology in Theory and Practice - Pattern and Process. Verlag New York: Springer.
Whittaker, R. H. (1956). Vegetation of the Great Smoky Mountains. Ecological Monographs, 26, 1-80.
Wu, J., Shen, W., Sun, W. & Tueller, P. T. (2002). Empirical pattern of the effects of changing scale on landscape metrics . Landscape Ecology, 17(8), 761–782.
Zuidema, P. A., Sayer, J. A. & Dijkman, W. (1996). Forest fragmentation and biodiversity: the case for intermediate-sized conservation areas . Environmental Conservation, 23(4), 290-297.
1.Biólogo, Doutorando no Programa de Pós-Graduação em Ciências Florestais, Universidade Federal Rural da Amazônia. ggaldino@gmail.com
2. Agronomia. Doutoranda no Programa de Pós-Graduação em Ciências Florestais, Universidade Federal Rural da Amazônia. marianaoliveirag22@hotmail.com
3. Bióloga, Professora do Instituto Socioambiental e dos Recursos Hídricos, Universidade Federal Rural da Amazônia. izildinha.miranda@ufra.edu.br
4. Bióloga, Pesquisadora do Institut de Recherche pour le Développement. danielle.mitja@ird.fr
6. Geógrafa, Pesquisadora do Institut de Recherche pour le Développement. anne-elisabeth.laques@ird.fr