![]() ISSN 0798 1015
ISSN 0798 1015

![]() ISSN 0798 1015
ISSN 0798 1015
Vol. 38 (Nº 44) Año 2017. Pág. 19
Rosinaldo de Sousa FERREIRA 1; Lucimara Ferreira de FIGUEREDO 2; Francisco Hélio Alves de ANDRADE 3; Sebastião de Oliveira de MAIA JÚNIOR 4; Rafael Ramos de MORAIS 5; Juarez Paz PEDROZA 6
Recibido: 10/05/2017 • Aprobado: 10/06/2017
RESUMO: Objetivou-se analisar a influência da torta de mamona e níquel no comportamento fisiológico da mamoneira. O delineamento experimental inteiramente casualizado, em esquema fatorial 4 x 4, sendo quatro doses de níquel (0,0; 4,0; 8,0; 12 mL da solução na concentração de 50 mg L-1) aplicadas via foliar e quatro doses de torta de mamona (0,0; 1,0; 2,0 e 3,0 t ha-1) com quatro repetições. Os dados foram submetidos à análise de variância e regressão. A torta de mamona exerceu efeito significativo sobre a eficiência quântica do fotossistema II, índice relativo de clorofila e trocas gasosas. Por sua vez, a aplicação de torta de mamona reduziu a transpiração, condutância estomática, eficiência instantânea do uso da água e a eficiência instantânea de carboxilação. |
ABSTRACT: The objective of this study was to analyze an influence of the castor bean cake and not a physiological behavior of the castor bean. The experimental design was completely randomized, in a 4 x 4 factorial scheme, with four doses of nickel (0.0, 4.0, 8.0, 12 mL of the solution in 50 mg L-1 concentration) applied by foliar and four doses Castor cake (0.0, 1.0, 2.0 and 3.0 t ha-1) with four replicates. Data were submitted to analysis of variance and regression. Castor cake had a significant effect on a quantum efficiency of photosystem II, relative index of chlorophyll and gas exchange. In turn, a castor bean application reduced transpiration, stomatal conductance, the instantaneous use of water and an instantaneous instance of carboxylation. |

A cultura da mamoneira no Brasil, destaca se o Nordeste é a principal região produtora, com destaque para os estados da Bahia e Ceará onde se localizam as maiores áreas cultivadas (LIMA et al., 2008), além de ter boa adaptabilidade, a mamona adquiriu relativa tolerância à seca (MARINHO et al., 2010).
Apesar de ser considerada uma planta de elevada resistência à seca, a mamona para produzir bem necessita de pelo menos 16 nutrientes essenciais (KIEHL, 1985) e aproximadamente 500 mm de precipitação pluvial bem distribuídos ao longo de seu ciclo, em especial na floração dos primeiros rasemos e solo de boa fertilidade natural.
A adubação é uma prática agrícola muito utilizada para a melhoria das culturas, atuando no fornecimento de nutrientes, melhorando as propriedades químicas e físicas do solo, e no fornecimento de nutrientes às culturas, na retenção de cátions (SEVERINO et al., 2006). Contudo, a definição de doses de fertilizantes químicos ou orgânicos que proporcionem a máxima eficiência quanto ao crescimento, desenvolvimento e produtividade, ainda é limitada.
A torta de mamona além de apresentar quantidades significativas de N, P e K, favorece grandes melhorias nas propriedades solo, por apresentar um elevado valor proteico encontrado e atraente ao pequeno produtor, além disto, é uma grande fonte de nitrogênio essencial para o desenvolvimento das plantas. De acordo com Lima et al. (2008) o uso da torta de mamona para o crescimento das plantas deve ser criteriosamente balanceado, visto que trata-se de uma fonte de matéria orgânica rica em nitrogênio, capaz de provocar danos irreversíveis e inviabilizar sua prática (ZUCHI et al., 2007).
Além do uso da adubação orgânica vem-se utilizando fertilizantes mineral via foliar contendo macro e micronutriente o níquel (Ni) é o elemento mais recentemente identificado como essencial para as plantas superiores (BROWN; WELSH; CARY, 1987); ocorre nos solos em várias formas: na solução do solo, trocável e não trocável, em minerais, e associado à matéria orgânica, por fazer parte da metaloenzima uréase, a qual participa da decomposição da uréia para amônia de CO2, deste modo, este elemento é importante para as plantas que recebem adubações com uréia ou seus derivados, exercendo papel importante no metabolismo de N (DECHEN; NACTTIGALL, 2007). A deficiência de Ni, impedindo a ação da uréase, leva ao acúmulo da uréia, o que causa manchas necróticas nas folhas. No mercado nacional é comercializado o nitrato de níquel, sem o embasamento científico para se justificar o uso crescente do produto em adubação foliar. Objetivou-se com esta pesquisa estudar os efeitos da torta de mamona e níquel no comportamento fisiológico da mamoneira BRS Energia.
O experimento foi conduzido em casa de vegetação, no Centro Nacional de Pesquisa (CNPA/Embrapa), localizado no município de Campina Grande, PB, no período de 16 de janeiro de 2014. A semeadura foi realizada manualmente, utilizando-se sementes da cv. BRS Energia, tratadas com fungicida, fornecidas pela própria Embrapa Algodão, sendo semeadas quatro sementes por vaso e, após 15 (DAS), de emergência realizou-se desbaste das plântulas deixando apenas uma planta por unidade experimental. O delineamento experimental utilizado foi inteiramente casualizado com 4 repetição e quatro concentrações de nitrato de Níquel (0,0; 4,0; 8,0; 12 mg L1) e quatro doses de torta de mamona ( 0,0; 1,0; 2,0; e 3,0 t/há-1) e totalizando assim, 16 tratamentos e 64 unidades experimentais.
No estudo cada unidade experimental foi representada por um vaso com capacidade de 30L, pintado de cor uniforme (prata), postos sobre o chão em fileiras espaçadas de 1,5 m e 1,0 m entre vasos por fileira. Posteriormente realizou-se incorporação da torta de mamona, ao solo a ser aplicado para cada unidade experimental foram ajustados proporcionalmente à quantidade de solo de cada.
Como substrato utilizou-se material de solo neossolo regolítico de textura franco-arenosa e torta de mamona (farelo) produzida com extração por solvente (fornecida pela indústria Brasil Óleo de Mamona, Salvador, BA). O resultados da análise da torta de mamona realizada no laboratório de análises químicas, pertencente a Embrapa Algodão segundo (COSTA et al., 2004), referentes ao nitrogênio (n), fósforo (p) e potássio (k), a torta contém 4,40 % de nitrogênio, 3,00 % de fósforo e 0,99 % de potássio.
As irrigações foram realizadas diariamente em quantidades iguais para todas as plantas, de acordo com as necessidades hídricas da cultura próximo a capacidade de campo.
Aos 20 dias após emergência da mamoneira foi realizadas aplicação via foliar as doses de Ni, por via foliar, por meio de um borrifador manual, visando a molhar as folhas completamente, mas evitando o escorrimento em intervalo de 15(DAS) totalizando três aplicações no decorre do ciclo.
Avaliaram-se as variáveis: fluorescência da clorofila ‘a’ (eficiência quântica do fotossistema II; fluorescência inicial; fluorescência máxima e fluorescência variável), nas medições utilizou-se o Fluorômetro PEA II – Plant Efficiency Analyser; o Índice relativo de clorofila foi avaliado em folhas totalmente expandidas do terço superior da planta em três pontos distintos de cada folha obtendo-se a média por planta, utilizando o aparelho portátil SPAD (“Soil plant analysis development”) e trocas gasosas (taxa de transpiração; condutância estomática; concentração interna de carbono; eficiência instantânea do uso da água e eficiência instantânea de carboxilação), as trocas gasosas foram realizadas na terceira folha de todas as parcelas, através de um analisador de gás infravermelho (IRGA), portátil, de acordo com a metodologia descrita por Marenco et al. (2001). Os dados obtidos no estudo foram submetidos à análise de variância pelo teste F, até 5% de significância os respectivos modelos de regressão foram ajustados de acordo com o coeficiente de determinação, até 5% de significância (STORCK et al., 2000).
As doses de torta de mamona promoveu efeito linear sobre a eficiência quântica do fotossistema II (Figura 1). Percebe-se que, as plantas de mamoneira sob a dose de 3 t ha-1 tornou-se mais eficiente proporcionando aumento de 33,12% comparado com a testemunha (0 t ha-1).
Santos et al. (2010), estudando atividade fotossintética em alface sob influência de diferentes doses de adubos orgânicos e fertilizantes minerais, os quais constataram para a eficiência quântica do fotossistema II em alface não foi alterada em função das diferentes doses de compostos orgânicos e minerais verificando maiores médias de 0,70 a 0,72, valores estes inferiores aos encontrados nesta pesquisa, sendo verificados valores médios entre 0,79 a 0,81.
Segundo Silva et al. (2007), plantas que estão com aparelho fotossintético intacto, apresentam valores médios da relação Fv/Fm variando de 0,75 a 0,85, enquanto que, reduções desses valores refletem danos fotoxidativos causados por fotoinibição no centro de reação do fotossistema II (PSII). As medições da eficiência fotoquímica do fotossistema II, estimadas através da relação entre a fluorescência variável e a fluorescência máxima da clorofila ‘a’ (Fv/Fm), relacionam-se com a eficiência quântica da atividade fotoquímica do PSII quando todos os centros de reação estão abertos (CORREIA et al., 2009). De acordo com Azevedo Neto et al. (2011) quando as plantas são expostas à luz, os centros de reação do PSII são progressivamente reduzidos, ocorrendo um aumento da fluorescência da clorofila, em seguida, a fluorescência decai em um fenômeno chamado extinção da fluorescência (“quenching”).
Por sua vez, a torta de mamona possui concentrações significativas de nitrogênio, essa concentração tem relação direta com a eficiência fotossintética. Segundo Larcher (2000), isto se deve ao fato das clorofilas absorverem quantidades de luz e serem responsáveis pela conversão desta energia luminosa à química, sendo mediada por fotorreceptores específicos. De acordo com Smeal e Zhang (1994); Larcher (2000), o processo acontece devido ao metabolismo dos cloroplastos decorrerem da incorporação deste elemento nas folhas pela dependência de enzimas, formadas basicamente de proteínas, ocorrendo assim, uma interdependência com o nitrogênio.
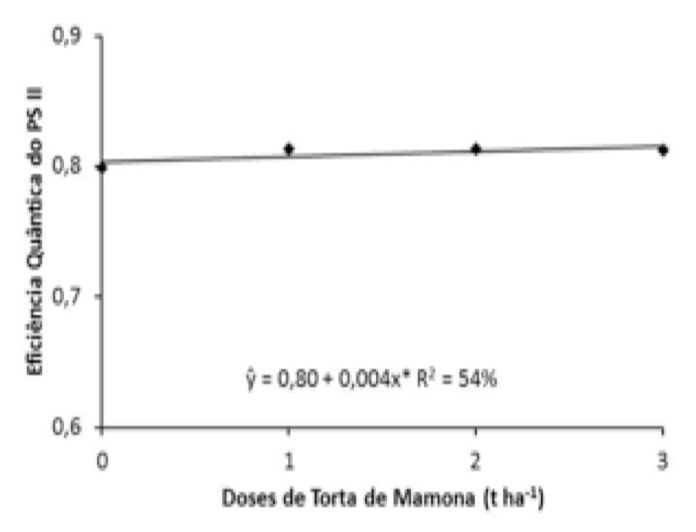
Figura 1. Eficiência quântica do fotossistema II (Fv/Fm)
em função de doses de torta de mamona.
Houve aumento linear do índice de clorofila das folhas de mamoneira em função da aplicação da torta de mamona, sendo que a maior dose (3,0 t ha-1) promoveu o valor máximo (53,37), atingindo incrementos de 9,69 %; 2,40 % e 0,69 %, da menor para maior dose analisada, respectivamente (Figura 2). De acordo com a tabela 2 pode-se observar que as doses níquel não exerceram influência sobre o índice de clorofila, e os resultados médios variaram de 51,16 a 53,28, mesmo não havendo, efeitos a maior dose (12,0 mL) aplicada propiciou o máximo resultado, este valor é inferior quando comparados com as plantas de mamoneira tratadas com torta de mamona, em uma porcentagem na ordem de 0,84 %. Estudando adubação orgânica em plantas de mamoneira Costa et al. (2011), também constataram um maior teor de clorofila nas maiores doses de torta de mamona aplicadas. Estes resultados eram aguardados e, pode ser explicado em função da melhoria das características físicas e químicas do solo fornecidos pela torta de mamona.
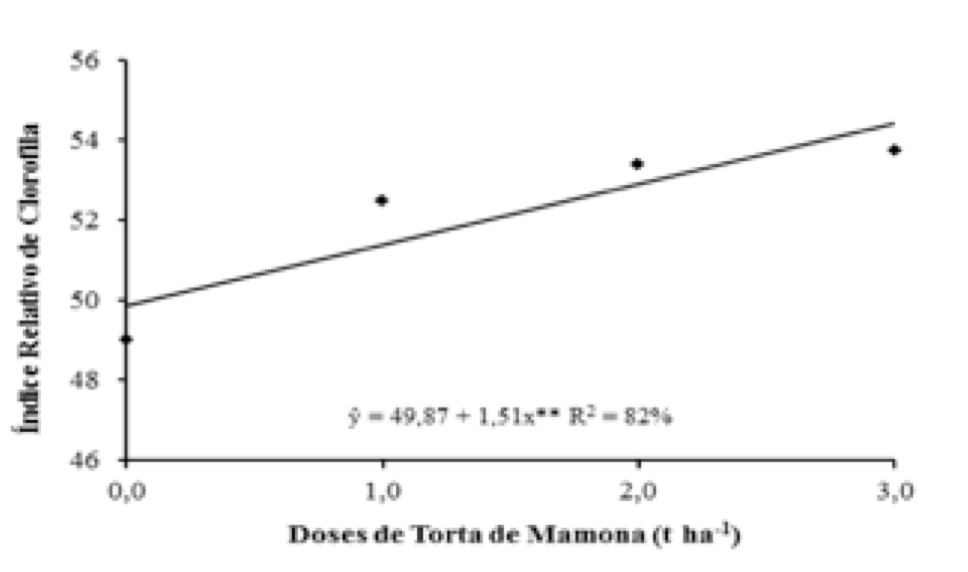
Figura 2. Índice relativo de clorofila em função de doses
de torta de mamona, em plantas de mamoneira.
Em relação aos efeitos da torta de mamona sobre a concentração interna de CO2 (Ci) verifica-se na figura 3 (A), que a dose inicial, ou seja, a testemunha propiciou a maior média observada (186,81 µmol mol-1), averiguando uma estabilidade nas doses intermediárias de 1,0 e 2,0 (t ha-1) e um aumento na dose máxima 3,0 t ha-1 (182,18 µmol mol-1). Assimilação de CO2 pelas folhas é reduzida principalmente pelo fechamento dos estômatos, onde ocorrem danos de membrana e redução da atividade enzimática, especialmente as de fixação de CO2 e a síntese de ATP (TAIZ; ZEIGER, 2009).
As doses de níquel apresentaram influência positiva para a concentração interna de CO2, havendo redução desta variável até a dose estimada 4,72 mL (182,12 µmol mol-1) e a dose de 4,0 mL o menor desempenho (163,18 µmol mol-1), (Figura 3 B). Machado et al. (1999) sugerem que o aumento de Ci, pode estar relacionado à queda na atividade de enzimas envolvidas no processo de fixação de CO2.
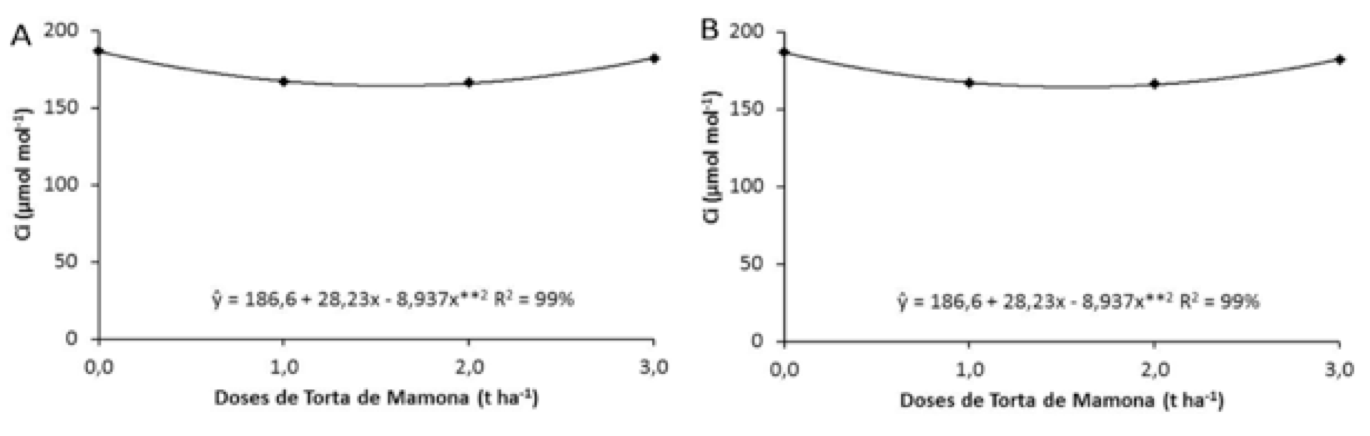
Figura 3. Concentração interna de CO2 (CI) em função de doses de torta
de mamona (A) e doses de níquel (B), sobre plantas de mamoneira.
A interação torta de mamona com níquel influenciou efeito sobre a taxa de transpiração (Figura 4A). Em virtude da dose de 3 t ha-1 de torta na presença de 8 mL de níquel foi observado maior taxa de transpiração em plantas de mamoneira.
Estudando o desdobramento do níquel dentro das doses de torta de mamona, constatou-se que o níquel obteve influência apenas sobre a dose de torta de 1,0 t ha-1, verificando aumento linear da taxa de transpiração com a elevação em uma unidade de níquel, obtendo-se um acréscimo de 24,54 %; 13,71 % e 9,85 %, da menor para maior dose de níquel aplicada (Figura 4B).
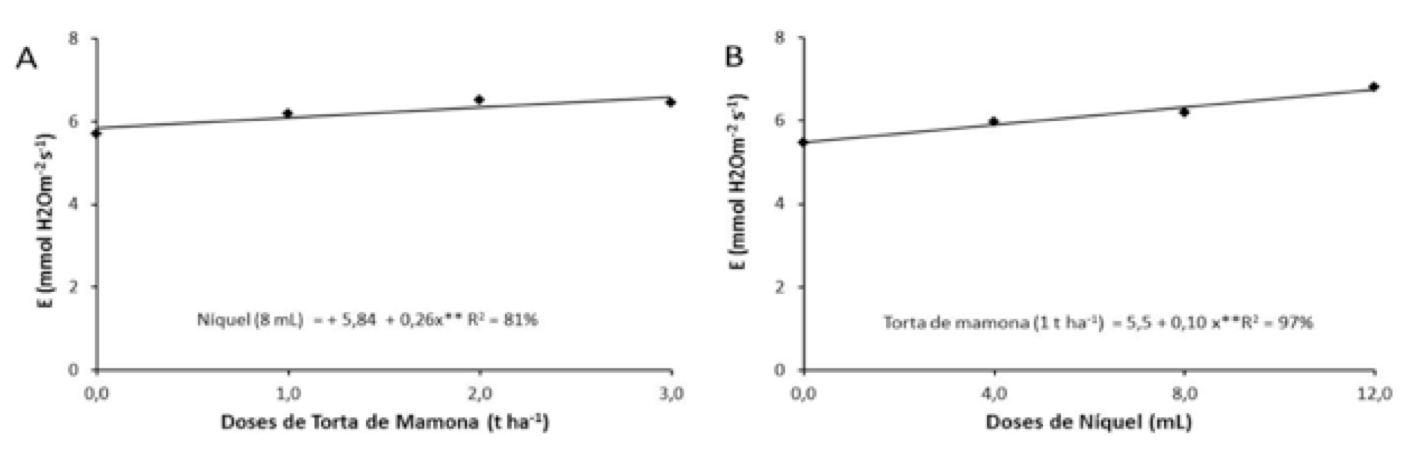
Figura 4. Desdobramento da taxa de transpiração (E) da torta de
mamona dentro da codificação de 8 mL de níquel (A), e Desdobramento
de níquel dentro da codificação de 1 t ha-1 de torta de mamona (B).
Estes resultados discordam com os encontrados por Santos et al. (2010), os quais não verificaram alteração das doses de adubação orgânica sobre a transpiração da cultura do alface. Segundo Machado et al. (2005) a transpiração é o principal meio envolvido na regulação da temperatura foliar e que menores aberturas estomáticas levam a redução da transpiração e aumento da temperatura do tecido foliar. Carvalho (2010) relata que o controle estomático da transpiração, atribui limitações difusivas ao CO2 que podem levar a um decréscimo na taxa fotossintética.
Verificando-se os efeitos de torta de mamona sobre a condutância estomática (Gs) apresentados na Figura 5, observa-se que a testemunha proporcionou o melhor resultado (0,412 mol m-2 s-1), apresentando redução de 15,73 %; 27,55 % e 13,46 %, respectivamente, da menor para maior dose. No entanto, as doses de níquel não exerceram efeitos sobre Gs, atingindo valores médios de 0,348 a 0,388 mols m-2 s-1, estes resultados são inferiores quando comparados com os obtidos pela torta de mamona, observou-se que a medida que se elevou uma unidade nas doses de torta ocorria um decréscimo na condutância estomática. Gs é um dos principais fatores que regulam o processo fotossintético nas plantas (PIMENTEL, 2004).
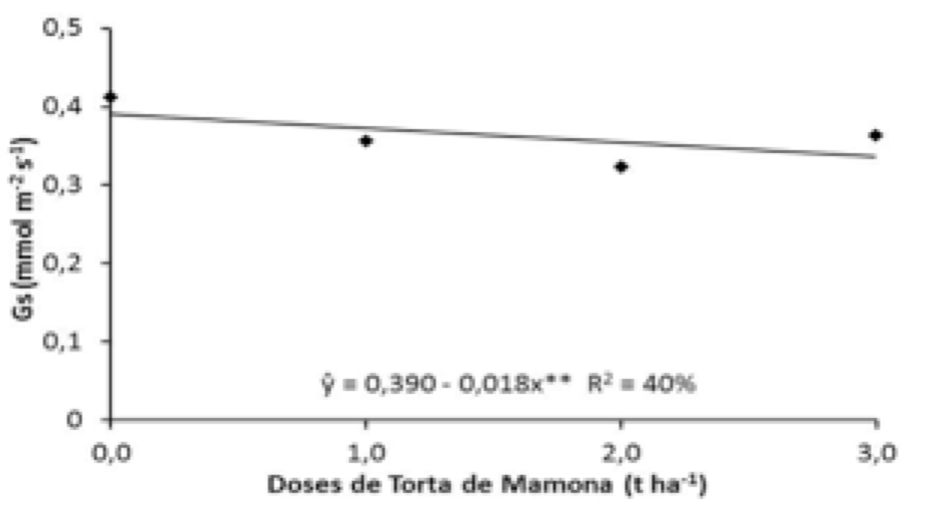
Figura 5. Condutância estomática (Gs) de plantas de
mamoneira em função de doses de torta de mamona.
Analisando-se o desdobramento da interação (T x N) sobre a eficiência instantânea do uso da água (A/E) (Figura 6 A), observa-se que a torta de mamona promoveu influência sobre a dose de níquel de 8,0 mL verificando redução na eficiência do uso da água com o aumento das doses de torta, com decréscimo de 5,5%; 11,16 % e 16,74 %, referentes à dose de níquel de 8,0 mL. Observando-se o desdobramento do níquel sobre as doses de torta de mamona (Figura 6 B), verifica-se que o níquel propiciou influência na dose de torta de mamona de 1,0 t ha-1, analisa-se que ocorreu redução linear com valor máximo promovido pelas plantas não adubadas com níquel (testemunha), atingindo diminuição de 7,51 %, 20,71 % e 27,22 %, respectivamente.
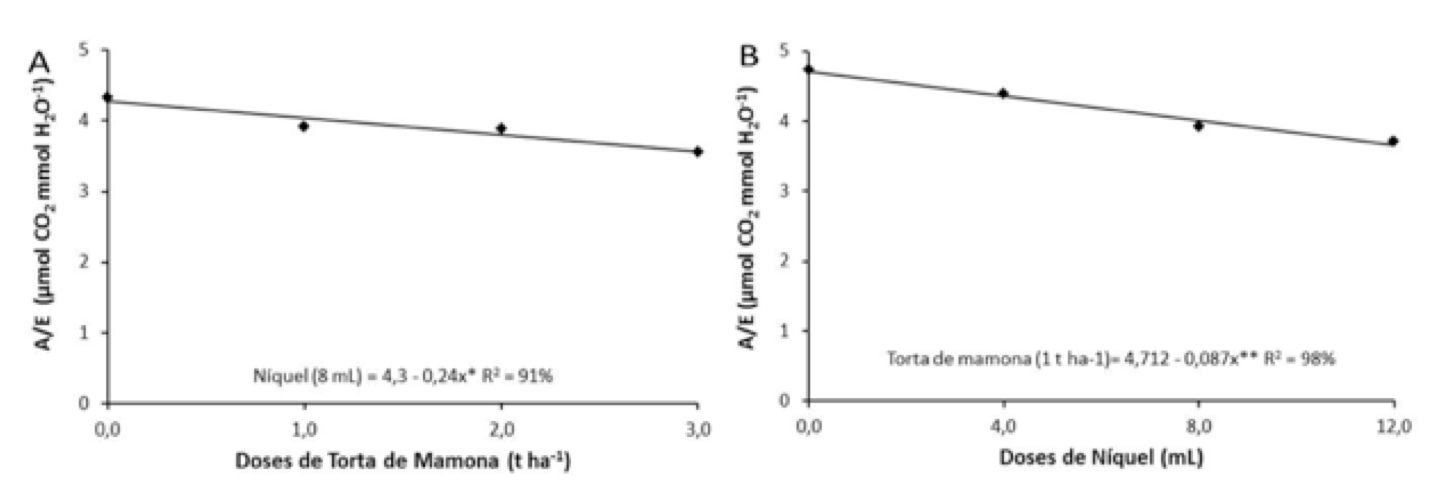
Figura 6. Desdobramento de torta de mamona dentro da dose de níquel (4 mL)
(A) e desdobramento de níquel dentro da dose de torta de mamona (1 t ha-1)
(B), sobre a eficiência instantânea do uso da água (A/E).
Resultados diferentes foram verificados nos estudos de Roza (2010) o qual relatou aumento nas razões A/E com o decréscimo da Gs. O autor reportou que o fechamento dos estômatos contribui para aperfeiçoar a eficiência no uso da água. Estudos constataram que em diferentes espécies cultivadas em ambiente controlado, com concentração de CO2 elevada, ocorreu aumento instantâneo da eficiência do uso da água, relacionado com a diminuição da condutância e transpiração (CANIZARES et al., 2004; ERISMANN et al., 2006).
Conforme observado na Figura 7 à análise de desdobramento sobre a eficiência instantânea de carboxilação (A/Ci), a torta de mamona exerceu influência dentro da dose de níquel de 8,0 mL, a maior eficiência de carboxilação foi proporcionada na dose de torta máxima estimada de 1,5 t ha-1. Estudo foram semelhantes aos encontrados por Carvalho (2010), o qual constatou baixa eficiência na carboxilação em plantas de mamoneira submetidas a estresse hídrico.
Diante dos resultados pode-se perceber que as plantas de mamona BRS Energia com torta de mamona para a variável A/Ci são sensíveis a altas concentrações de torta de mamona evidenciando uma queda na eficiência da carboxilação. Valores elevados de concentração interna de CO2 associado ao aumento na condutância estomática indicam um acréscimo na eficiência de carboxilação, ocorrido em função da disponibilidade de ATP e NADPH e do substrato para a Rubisco. Sendo assim, a A/Ci depende da disponibilidade de CO2 no mesófilo foliar, quantidade de luz, temperatura e da atividade enzimática para que ocorra a fotossíntese. Se as concentrações de CO2 intercelulares são muito baixas, o influxo deste componente nas células do mesófilo é restringido, assim, a planta aproveita o CO2 originário da respiração para cultivar um coeficiente mínimo de taxa fotossintética, tornando-a limitada (SALAMONI, 2010).
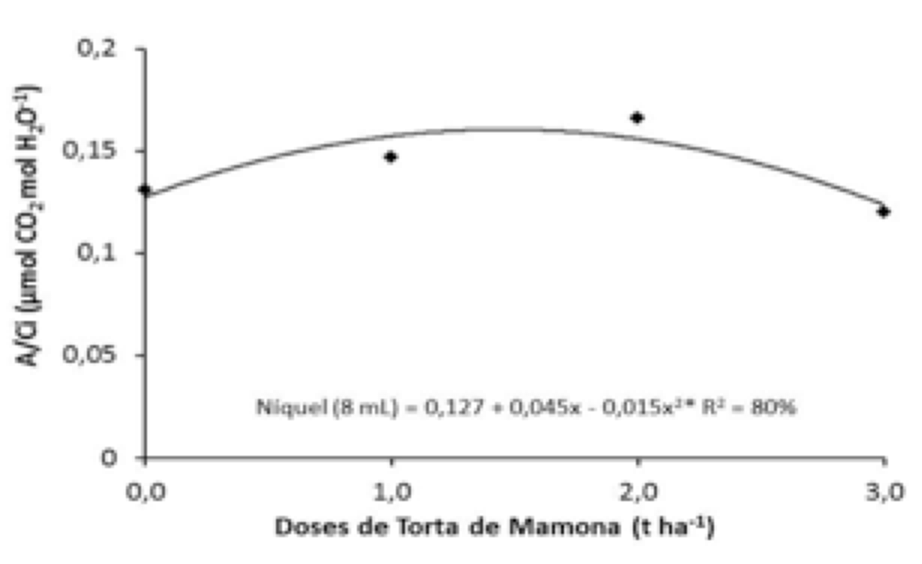
Figura 7. Desdobramento de torta de mamona dentro da dose de níquel (8 mL)
sobre a Eficiência Instantânea de Carboxilação (A/Ci).
Não houve alteração na eficiência quântica do fotossistema II e no índice relativo de clorofila em resposta a aplicação de níquel;
A torta de mamona exerceu influencia sobre a fluorescência do fotossistema II de 0,812;
A aplicação de torta de mamona reduziu a condutância estomática, eficiência instantânea do uso da água;
A dose de 1,5 t ha-1 promoveu maior eficiência instantânea de carboxilação de 0,163.
AZEVEDO NETO, A. D. et al. Fluorescência da clorofila como uma ferramenta possível para seleção de tolerância à salinidade em girassol. Revista Ciência Agronômica, Fortaleza, v. 42, n. 4, p. 893-897, 2011.
BROWN, P.H.; WELSH, R.M.; CARY, E.E. Nickel: A micronutrient essential forhigher plants. Plant Physiology, Rockville, v.85, n.3, p.801-803, 1987.
CANIZARES, K. A. L.; RODRIGUES, J. D.; GOTO, R. Crescimento e índices de troca gasosa em plantas de pepino irrigadas com água enriquecida com CO2. Horticultura Brasileira, Brasília, v. 22, n.4, p. 706-711, 2004.
CARVALHO, M. Variações morfofisiológicas e eficiência do uso da água em Ricinus communis L. submetida ao déficit hídrico regulado. Ilhéus, 2010. 106p. Dissertação (Mestrado em Produção Vegetal). Programa de Pós – Graduação em Produção Vegetal, Universidade Estadual de Santa Cruz, Ilhéus, 2010.
CORREIA, K. G. et al. Crescimento, produção e características de fluorescência da clorofila a em amendoim sob condições de salinidade. Revista Ciência Agronômica, Fortaleza, v. 40, n. 4, p. 514 – 521, 2009.
COSTA, F. X. et al. Avaliação da fisiologia e bioquímica da mamoneira em função da aplicação de composto orgânico de lixo e torta de mamona como fertilizantes. Revista Engenharia Ambiental, Espiríto Santo do Pinhal, v. 8, n.1, p. 101-109, 2011.
COSTA, K.A.P.; FRANÇA, A.F.S.; OLIVEIRA, I.P. et al. Composição química-bromatológica do capim-tanzânia em função de doses de nitrogênio, potássio e enxofre. Cienc. Anim. Bras. , v.25, p.83-91, 2004.
DECHEN, A. R.; NACTTIGALL, G. R. Elementos requeridos à nutrição de plantas. In: NOVAIS, R. F.; ALVAREZ, V. V.; BARROS, N. F.; FONTES, R. L. F.; CANTARUTT, R. B.; NEVES, J. C. L. (ed). Fertilidade do Solo. Viçosa. Sociedade Brasileira de Ciências do Solo, p. 91- 132, 2007.
ERISMANN, N. M.; MACHADO, E. C.; GODOY, I. J. Capacidade fotossintética de genótipos de amendoim em ambiente natural e controlado Pesquisa Agropecuária Brasileira, v. 41, n. 7, p. 1099-1108, 2006.
KIEHL, E.J. Fertilizantes orgânicos. Piracicaba, Editora Agronômica Ceres Ltda, 1985. 492p
LARCHER, W. Ecofisiologia vegetal. São Carlos, Rima, 2000. 531p.
LIMA, R. L. S. et al. Casca e torta de mamona avaliados em vasos como fertilizantes orgânicos. Revista Caatinga, Mossoró, v. 21 n. 5 (Número especial), p.102-106, dez. 2008.
MACHADO, E. C. et al. Respostas da fotossíntese de três espécies de citros a fatores ambientais. Pesquisa Agropecuária Brasileira, Brasília, v. 40, n. 12, p. 1161-1170, 2005.
MACHADO, E. C., MEDINA, C. L., GOMES, M. M. A. Teor de água no substrato de crescimento e fotossíntese em laranjeira “Valença”. Bragantia, Campinas, v. 58, n. 1, p. 217-226, 1999.
MARENCO, R. A.; GONÇALVES, J. F. C.; VIEIRA, G. Leaf gás Exchange and carbohydrates in tropical tress differing in successional status in two light environments in Central Amazonia. Tree Physiology, v. 21, n. 18, p. 1311-1318, 2001.
MARINHO, A. B. et al. Influência da fertirrigação da nitrogenada na produtividade da cultura da mamoneira. Revista Brasileira de Agricultura Irrigada, v. 4, n. 1, p. 31-42, 2010.
PIMENTEL, C. A relação da planta com a água. Rio de Janeiro: Edur, 2004, 191p.
ROZA, F. A. Alterações morfofisiológicas e eficiência de uso da água em plantas Jatropha curcas L. submetidas à deficiência hídrica. Ilhéus, 2010, 78f. Dissertação (Mestrado em Produção Vegetal). Universidade Estadual de Santa Cruz, Ilhéus, 2010.
SALAMONI, A. T. Fotossíntese – Ecofisiologia. In: Apostila de aulas teóricas e práticas de fisiologia vegetal. Santa Maria, RS. Cap. VII, p. 28-29, 2010.
SANTOS, C. M. et al. Atividade fotossintética em alface (Lactua sativa L.) submetidas a diferentes compostagens de resíduos agroindustriais. Pesquisa Aplicada & Agrotecnologia, Guarapuava, v. 3, n. 3, p. 103-112, 2010.
SANTOS, C. M.; GONÇALVES, E. R.; ENDRES, L.; GOMES, T. C. A.; JADOSKI, C. J.; NASCIMENTO, L. A.; SANTOS, E. D. Atividade fotossintética em alface (Lactuca sativa L.) submetidas a diferentes compostagens de resíduos agroindustriais. Pesquisa Aplicada & Agrotecnologia, v.3, n.3, p. 95-102, 2010.
SEVERINO, L. S.; MILANI, M.; BELTRÃO, N. E. M. Mamona: O produtor pergunta, a Embrapa responde. Brasília, DF: Embrapa Informação Tecnológica; Campina Grande: Embrapa Algodão, 2006. (Coleção 500 perguntas, 500 respostas).
SILVA, T. R. B. et al. Adubação nitrogenada em cobertura na cultura da mamona em plantio direto. Pesquisa Agropecuária Brasileira, Brasília, v. 42, n. 9, p. 1357-1359, 2007.
SMEAL, D; ZHANG, H. Chlorophyll meter evaluation for nitrogen management in corn. Communications in Soil Science and Plant Analysis, New York, v. 25, n. 9-10, p.1495-1503, 1994.
STORCK, G. D. C.; LOPES, S. J.; ESTEFANEL, V. Experimental agrícola. Santa Maria: Editoria da UFSM, 2000.
TAIZ, L.; ZEIGER, E. Fisiologia Vegetal. 4. ed. Porto Alegre: Artmed, 2013. 848p.
ZUCHI, J. et al. Efeito de torta de mamona sobre componentes de rendimento de trigo. Revista Brasileira de Agroecologia, Fortaleza, v. 2, n. 2, p. 1080 –1083, 2007.
1. Programa de Pós - Graduação em Agronomia, Universidade Federal da Paraíba, doutorando em agronomia. Email: rosinaldoagrarias@hotmail.com
2. Programa de Pós - Graduação em Agronomia, Universidade Federal da Paraíba, doutoranda em agronomia. Email: lucimara.ufpb@gmail.com
3. Programa de Pós - Graduação em Agronomia, Universidade Federal da Paraíba ,mestrando em agronomia. Email: helioalvesuepb@gamil.com
4. Programa de Pós - Graduação em Agronomia, Universidade Federal de Alagoas, doutorando em agronomia. Email: juniormaiagrari@hotmal.com
5. Programa de Pós - Graduação em Agronomia, Universidade Federal da Paraíba, mestrando em agronomia. Email: leafarmorais@gmail.com
6. Prof. Dr. da Universidade Federal de Campina Grande/UFCG. Email: juarez@deag.ufcg.edu.br